the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 03 Aug 2022
| 03 Aug 2022
Changes in productivity and intermediate circulation in the northern Indian Ocean since the last deglaciation: new insights from benthic foraminiferal Cd ∕ Ca records and benthic assemblage analyses
Ruifang Ma
Sophie Sépulcre
Laetitia Licari
Frédéric Haurine
Franck Bassinot
Zhaojie Yu
Christophe Colin
We have measured ratios of several benthic foraminiferal species and studied benthic foraminiferal assemblages on two cores from the northern Indian Ocean (Arabian Sea and northern Bay of Bengal, BoB), in order to reconstruct variations in intermediate-water circulation and paleo-nutrient content since the last deglaciation. Intermediate water Cdw records estimated from the benthic reflect past changes in surface productivity and/or intermediate–bottom-water ventilation. The benthic foraminiferal assemblages are consistent with the geochemical data. These results suggest that during the last deglaciation, Cdw variability was primarily driven by changes in intermediate-water properties, indicating an enhanced ventilation of intermediate–bottom water masses during both Heinrich Stadial 1 and the Younger Dryas (HS1 and YD, respectively). During the Holocene, however, surface primary productivity appears to have influenced Cdw more than intermediate water mass properties. This is evident during the early Holocene (from 10 to 6 cal ka) when benthic foraminiferal assemblages indicate that surface primary productivity was low, resulting in low intermediate-water Cdw at both sites. Then, from ∼ 5.2 to 2.4 cal ka, surface productivity increased markedly, causing a significant increase in the intermediate-water Cdw in the southeastern Arabian Sea and the northeastern BoB. The comparison of intermediate-water Cdw records with previous reconstructions of past Indian monsoon evolution during the Holocene suggests a direct control of intermediate-water Cdw by monsoon-induced changes in upper-water stratification and surface primary productivity.
- Article
(16891 KB) - Full-text XML
-
Supplement
(1105 KB) - BibTeX
- EndNote





During the last deglaciation, a two-step rapid increase in atmospheric CO2 occurred during the 17–13.8 and 12.3–11.2 cal ka time intervals (e.g., Monnin et al., 2001). Several studies suggest that variations in the Southern Ocean circulation contributed to these increases in atmospheric CO2 by transferring deep-ocean carbon to the upper ocean and atmosphere, through enhanced upwelling and increased northward penetration of the Antarctic Intermediate Water (AAIW) in all ocean basins (e.g., Marchitto et al., 2007; Anderson et al., 2009; Skinner et al., 2014). Different proxies have been used to reconstruct past changes in intermediate circulation, such as radiocarbon activity (Δ14C) (e.g., Marchitto et al., 2007; Bryan et al., 2010), benthic δ13C (e.g., Pahnke and Zahn, 2005; Jung et al., 2009; Ma et al., 2019), foraminiferal εNd (e.g., Pahnke et al., 2008; Xie et al., 2012; Yu et al., 2018) and benthic foraminifera (Ma et al., 2020). These studies have focused on the close relationship between enhanced ventilation in the Southern Ocean and rising atmospheric CO2 during the last deglaciation period. Furthermore, it has been shown that glacial–interglacial transfer of CO2 between the oceans and the atmosphere could also be linked to changes in the efficiency of the oceanic biological pump (Pichevin et al., 2009; Ziegler et al., 2013; Bauska et al., 2016; Hertzberg et al., 2016; Jaccard et al., 2016; Yu et al., 2019), which may contribute to up to half of the observed CO2 flux (Kohfeld et al., 2005).
The oceanic biological pump and nutrient upwelling are at least partly controlled by intermediate–deep-water circulation, contributing to the observed CO2 changes (e.g., Toggweiler, 1999; Marchitto and Broecker, 2006). To track past changes in the nutrient concentration of intermediate water masses, benthic foraminifera has been used in many recent studies (e.g., Came et al., 2008; Poggemann et al., 2017; Valley et al., 2017; Umling et al., 2018); indeed, the benthic foraminifera is a robust proxy for seawater cadmium concentrations (Cdw) (Boyle, 1988, 1992), which show a positive linear correlation with labile nutrients (phosphate and nitrate) in the modern ocean (e.g., Boyle et al., 1976; Boyle, 1988; Elderfield and Rickaby, 2000). The benthic foraminifera incorporate Cd as a function of Cdw with a species-dependent partition coefficient (e.g., Tachikawa and Elderfield, 2002). Thus, the Cd measured in the fossil tests reflects the paleo-nutrient concentrations of the surrounding water masses and can be used to investigate past changes in intermediate-to-deep-ocean properties (e.g., Boyle and Keigwin, 1982; Oppo and Fairbanks, 1987; Came et al., 2008; Poggemann et al., 2017; Valley et al., 2017; Umling et al., 2018).
Complementary to the geochemical proxies, the type of benthic foraminifers and their abundance, both of which are related to organic flux and ecosystem oxygenation, make benthic foraminifer assemblages a powerful proxy for estimating past variations in bottom-water conditions (e.g., Corliss et al., 1986; Schmiedl et al., 1998; Almogi-Labin et al., 2000) in conjunction with organic matter fluxes to the seafloor (e.g., Altenbach et al., 1999; Van der Zwaan et al., 1999; Fontanier et al., 2002; Caulle et al., 2015). Benthic foraminifera have been successfully used as indicators of surface productivity, especially in high-carbon-flux regions (Schnitker, 1994). By comparing past benthic foraminiferal assemblages to modern ones, changes in food supply and oxygen concentrations of the bottom water can be reconstructed (e.g., Corliss, 1979; Peterson, 1984; Murgese and De Deckker, 2005). Recently, the combining of benthic foraminiferal assemblages and geochemical proxies has received increasing attention and has been used to reconstruct the evolution of surface productivity and upwelling intensity in the Indian Ocean (e.g., Hermelin 1991, 1992; Hermelin and Shimmield, 1995; Den Dulk et al., 1998; Murgese and De Deckker, 2005).
The Arabian Sea is one of the most productive regions of the ocean today (Banse, 1987; Marra and Barber, 2005). Surface productivity is dominated by the monsoon system, which has a strong impact on the distribution and dynamics of stratification and vertical mixing (Lévy et al., 2007). Numerous studies have focused on the reconstruction of the paleo-productivity of the Arabian Sea in relation to past changes in monsoon intensity (e.g., Prell and Kutzbach, 1987; Naidu and Malmgren, 1996; Gupta et al., 2003; Singh et al., 2006, 2011; Bassinot et al., 2011; Saraswat et al., 2014). By contrast, little is known about the paleo-productivity of the Bay of Bengal (BoB), especially its links to changes in monsoon precipitation (Phillips et al., 2014; Zhou et al., 2020). Consequently, studying paleo-productivity and past nutrient concentration of intermediate water masses in the northeastern Indian Ocean will also allow us to completely understand the influence of monsoon climate changes in tropical ocean ecology at different timescales. Moreover, as the benthic foraminiferal is a promising proxy to reconstruct the intermediate–deep-water nutrient content (e.g., Boyle and Keigwin, 1982; Tachikawa and Elderfield, 2002; Came et al., 2008; Poggemann et al., 2017; Valley et al., 2017), most of the studies referred to above have reconstructed deep–intermediate water masses in the past (e.g., Came et al., 2008; Bryan and Marchitto, 2010; Poggemann et al., 2017; Valley et al., 2017), and only few works investigate the relationship between the intermediate water mass nutrient and surface productivity (Bostock et al., 2010; Olsen et al., 2016). Furthermore, the evolution of the nutrient content of intermediate water masses since the last deglaciation has never been reconstructed in the Indian Ocean, where only two low-resolution records are available for deep water depths (Boyle et al., 1995), and, to our knowledge, none are available for intermediate water depths.
In this study, we provide, for the first time, two benthic foraminifera records at intermediate water depths in the northern Indian Ocean (Arabian Sea and northern Bay of Bengal). These data make it possible to estimate past changes in the nutrient content since the last deglaciation. We have also investigated benthic foraminiferal assemblages obtained from core MD77-191 (southeastern Arabian Sea) to help us reconstruct the conditions at the seafloor. Combined with planktonic foraminiferal δ18O, benthic δ13C and records obtained from the same core as well as with results already published in the Bay of Bengal (Ma et al., 2019, 2020), this study aims to document past variations in intermediate- and deep-water conditions and to decipher their links with surface paleo-productivity and intermediate-water ventilation.
We analyzed sediment core MD77-191 (07∘30′ N, 76∘43′ E, 1254 m) located in the Arabian Sea (off the southern tip of India), and core MD77-176 (14∘30′ 5′′ N, 93∘07′6′′ E, 1375 m) retrieved in the northeastern Bay of Bengal (BoB). These cores were collected in 1977 during the OSIRIS III cruise of the French R/V Marion Dufresne (Fig. 1).
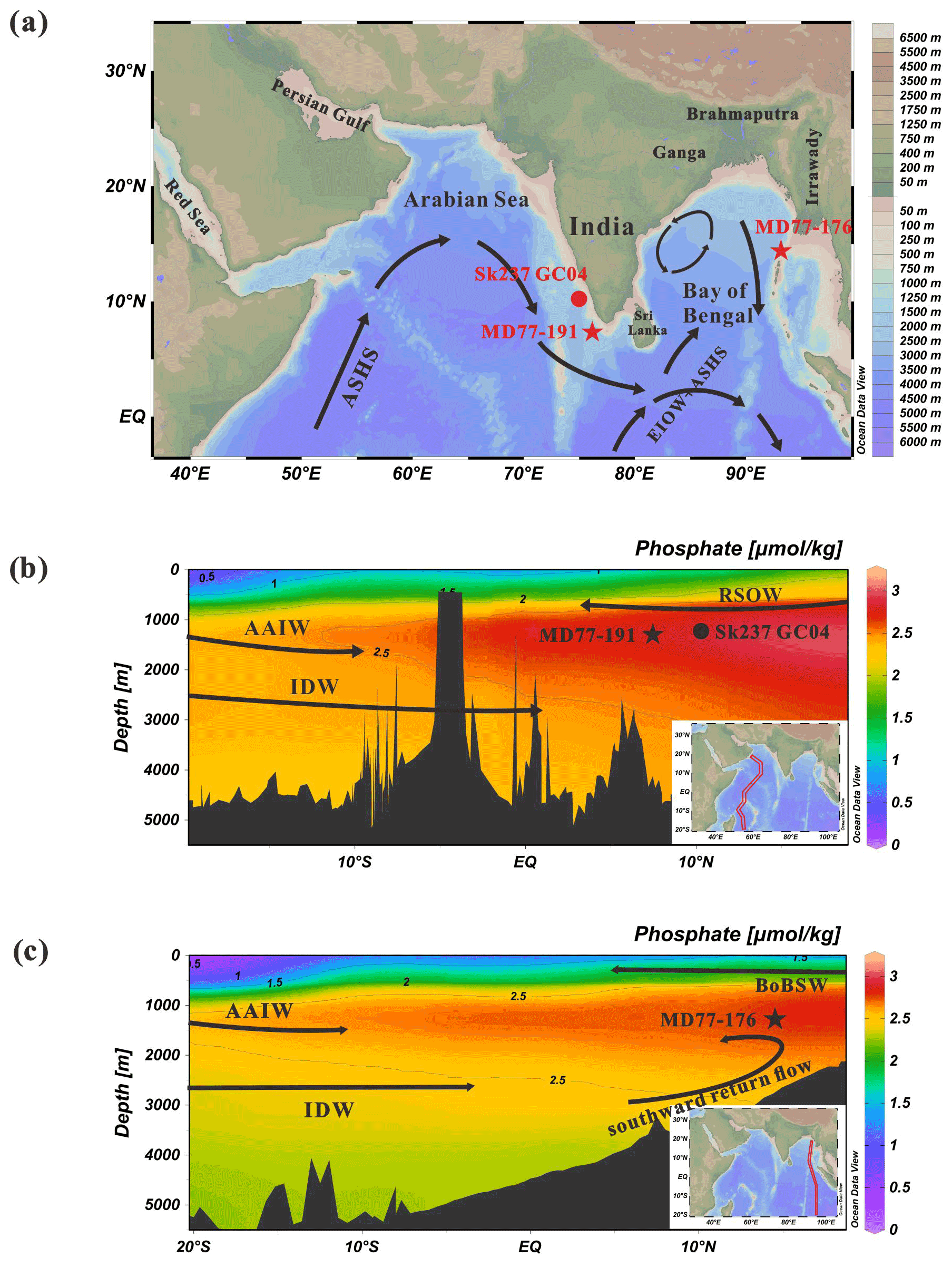
Figure 1(a) Oceanographic setting and locations of core MD77-191 in the Arabian Sea (red star), core MD77-176 in the Bay of Bengal (red star) and reference site SK237 GC04 (red circle, Naik et al., 2017). The black arrows represent the general surface circulation direction in the northern Indian Ocean during the summer southwest monsoon (Schott and McCreary, 2001). (b, c) Phosphate distribution along depth–latitude sections during the southwest monsoon and northeast monsoon periods, for the Arabian Sea and the Bay of Bengal, respectively. Data (in µmol kg−1, colored scale; Schlitzer, 2000) were contoured and plotted using the Ocean Data View (ODV) software (Schlitzer, 2015). On these two figures are shown the distribution and circulation of water masses in the Arabian Sea and Bay of Bengal (black arrows). ASHS: Arabian Sea High Salinity Water; EIOW: Eastern Indian Ocean Water; BoBSW: Bay of Bengal surface waters; AAIW: Antarctic Intermediate Water; RSOW: Red Sea Overflow Water; IDW: Indian Deep Water.
The age model of core MD77-191 was established by using accelerator mass spectrometry (AMS) 14C dates obtained on nine monospecific samples of planktonic foraminifera Globigerinoides bulloides (Bassinot et al., 2011), one sample of pteropods (Mléneck, 1997) and three samples of the planktonic foraminifera Globigerinoides ruber (Ma et al., 2020). The average sedimentation rate of core MD77-191 is about 53 cm kyr−1 and up to 90 cm kyr−1 during the Holocene, providing a high-resolution, continuous record since 17 cal kyr BP.
The age model of core MD77-176 was previously established by using 31 planktonic foraminifer (G. ruber) AMS 14C dates combined with the core MD77-176 oxygen isotope record obtained on planktonic foraminifer G. ruber, which were correlated to the GISP2 Greenland ice core record (Marzin et al., 2013). Core MD77-176 displays high accumulation rates (average ∼25 cm kyr−1 and up to 40 cm kyr−1 during the Holocene).
In the modern ocean, the surface waters of the Arabian Sea and BoB are characterized by seasonally reversing currents that are driven by the monsoon winds (Fig. 1a). The surface water masses shallower than 150 m in the Arabian Sea are mainly Arabian Sea High Salinity Water (ASHS, 36.5 psu) (Talley et al., 2011). In the BoB, the surface waters above 100 m are designated Bay of Bengal surface waters (BoBSW), which have a low salinity (31 psu) due to large river inputs (Talley et al., 2011). Today, the northward extension of AAIW in the Indian Ocean rarely reaches beyond 10∘ S (Lynch-Stieglitz et al., 1994). The sites of cores MD77-191 and MD77-176 are mainly bathed, therefore, by the North Indian Intermediate Water (Olson et al., 1993; Reid, 2003) with a potential contribution from the Red Sea Outflow Water (RSOW) for the site MD77-191 (Beal et al., 2000).
Due to the land–sea configuration in the north by Asia, the deep waters of the northern Indian Ocean originate from the south, including the Circumpolar Deep Water (CDW) and North Atlantic Deep Water (NADW) (You, 2000; Tomczak and Godfrey, 2003; Talley et al., 2011). Thus, between 1500 and 3800 m, the dominant deep water in the northern Indian Ocean is Indian Deep Water (IDW), originating from the CDW admixed with NADW (You, 2000; Tomczak and Godfrey, 2003; Talley et al., 2011). Then, on their pathway, the bottom water upwells when it expands northward in the northern Indian Ocean, returning to shallower depths (You, 2000; Fig. 1c). Therefore, variations in deep water masses can also influence the intermediate-depth waters in the northern Indian Ocean.
As far as surface waters are concerned, during the summer monsoon, the clockwise circulation in the Arabian Sea drives high-salinity waters from the northern to the southeastern Arabian Sea. By contrast, during the winter monsoon, the northeastern winds bring low-salinity water (BoBSW) from the BoB. The northern Indian Ocean, especially the Arabian Sea, is characterized by highly variable seasonal productivity (Shankar et al., 2002). Southwest winds during the summer season induce a strong Ekman pumping resulting in very active upwelling along the western coasts of the Arabian Sea and thus promoting strong surface productivity (Shankar et al., 2002; Fig. S1 in the Supplement). By contrast, the surface productivity in the BoB is generally weak compared with the Arabian Sea (e.g., Prasanna Kumar et al., 2001; Thushara and Vinayachandran, 2016; O'Malley, 2017; Fig. S1). In the BoB, large river inputs of freshwater and direct monsoon precipitation lead to more stable stratification in the upper ocean (Vinayachandran et al., 2002), and hence the vertical mixing of nutrients from the subsurface to the euphotic zone is generally limited (Gomes et al., 2000). However, the primary productivity of the western BoB shows a slight increase during the winter monsoon, as indicated by the distribution of chlorophyll in the surface water (Thushara and Vinayachandran, 2016; O'Malley, 2017; Fig. S1).
Modern data indicate that the southern-sourced intermediate water (AAIW) in the Indian Ocean has a phosphate concentration of about 2–2.5 µmol kg−1 (Fig. 1b and c). In the northern intermediate Indian Ocean, the phosphate concentration is significantly higher, ranging from 2.75 to 3 µmol kg−1 in the Arabian Sea during the summer monsoon, and from 2.5 to 2.75 µmol kg−1 in the BoB during the winter monsoon (Fig. 1b and c). The higher phosphate in the northern Indian Ocean can been linked to increased primary productivity (Banse, 1987; Marra and Barber, 2005).
3.1 Cd Ca analysis
In order to improve understanding of possible inter-species differences and microhabitat effects on the benthic records, we analyzed in three calcite (Cibicidoides pachyderma, Uvigerina peregrina and Globobulimina spp.) and one aragonite (Hoeglundina elegans) benthic foraminiferal species from core MD77-191. C. pachyderma is a shallow infaunal species U. peregrina and Globobulimina spp. are endobenthic species with intermediate and deep microhabitats, respectively (Fontanier et al., 2002). In core MD77-176, due to the limitation of calcitic species, we only measured ratios in H. elegans shells. Moreover, , and ratios were also measured in all benthic foraminiferal samples to check the robustness of results and the potential influence of contamination (i.e., oxides and sedimentary clay, Barker et al., 2003).
Each sample contained between 10 and 15 individuals picked from the 250–315 µm size fraction. Samples were gently crushed and cleaned to remove clays, organic matter and elemental oxides by using reductive and oxidative cleaning following previously published methods (Boyle and Keigwin, 1982; Barker et al., 2003). Each sample was dissolved in 0.075 N HNO3 and analyzed using a single collector sector field high-resolution inductively coupled plasma mass spectrometer (HR-ICP-MS) Thermo Element XR hosted at the GEOPS Laboratory (University Paris-Saclay, France).
The detailed instrumental settings and mother standard solutions are described in Ma et al. (2020). A blank consisting of the same 0.1 N HNO3 used to dilute the standards and samples was also analyzed. We removed the blank intensity values from all the raw intensities (including standards), and raw data were linearly drift-corrected by interspersing a drift standard every four samples. Standard curves were used to calculate elemental concentrations, coefficients of determination (r2) always being >0.9999 for all elemental ratios. The mean reproducibility and accuracy are 3.6 % and 7.5 %, respectively.
3.2 Faunal analysis
Benthic foraminiferal assemblages from core MD77-176 have already been published in Ma et al. (2019). For core MD77-191, a total of 72 samples were collected for benthic foraminiferal assemblage determinations. In each sample, benthic foraminifera (>150 µm) were extracted, counted and identified to species level following the taxonomical descriptions of various authors (e.g., Loeblich and Tappan, 1988; Jones, 1994; Holbourn et al., 2013). For core MD77-191, there is no material left in this old, small-diameter core, and so we used samples obtained earlier for stable isotope studies. Since the bulk weights of these samples were not recorded prior to sieving, we could not perform the calculation of the absolute abundance of foraminifera or accumulation rates. Thus, we only converted the individual counts to percentages with respect to the total benthic foraminifera present in each sample. In order to describe major faunal variations, we performed principal component analysis (PCA) on the variance–covariance matrix using the PAST software (Version 3.0, Hammer et al., 2001). Species present with a percentage >1 % in at least 1 sample were used for statistical analysis and diversity calculation.
4.1 Elemental ratio results
To check the influence of oxide contaminants on the elemental ratios, was systematically measured. The of H. elegans from cores MD77-191 and MD77-176 ranges between 6.5–10 and 1–30 µmol mol−1, respectively. Such ranges are much lower than the 100 µmol mol−1 limit proposed by Boyle (1983). The values obtained on the three calcite benthic foraminifera species from core MD77-191 – C. pachyderma (5–18 µmol mol−1), U. peregrina (3–23 µmol mol−1) and Globobulimina spp. (4–69 µmol mol−1) – are also all below 100 µmol mol−1 (Boyle, 1983). The ratios are also lower than 1 mmol mol−1 in all samples from cores MD77-191 and MD77-176, in agreement with the limit proposed by Barker et al. (2003). In addition, Barker et al. (2003) concluded that no significant pollution by clay minerals would be expected when is <0.5 mmol mol−1. In all our samples, is below 0.5 mmol mol−1, indicating that the sample cleaning procedure was efficient.
All of the above results indicate that our samples were not affected by contamination.
Cd Ca
The records of C. pachyderma, U. peregrina and Globobulimina spp. from core MD77-191 range between 0.07–0.2, 0.07–0.14 and 0.03–0.09 µmol mol−1, respectively (Fig. 2d; Table S1 in the Supplement).
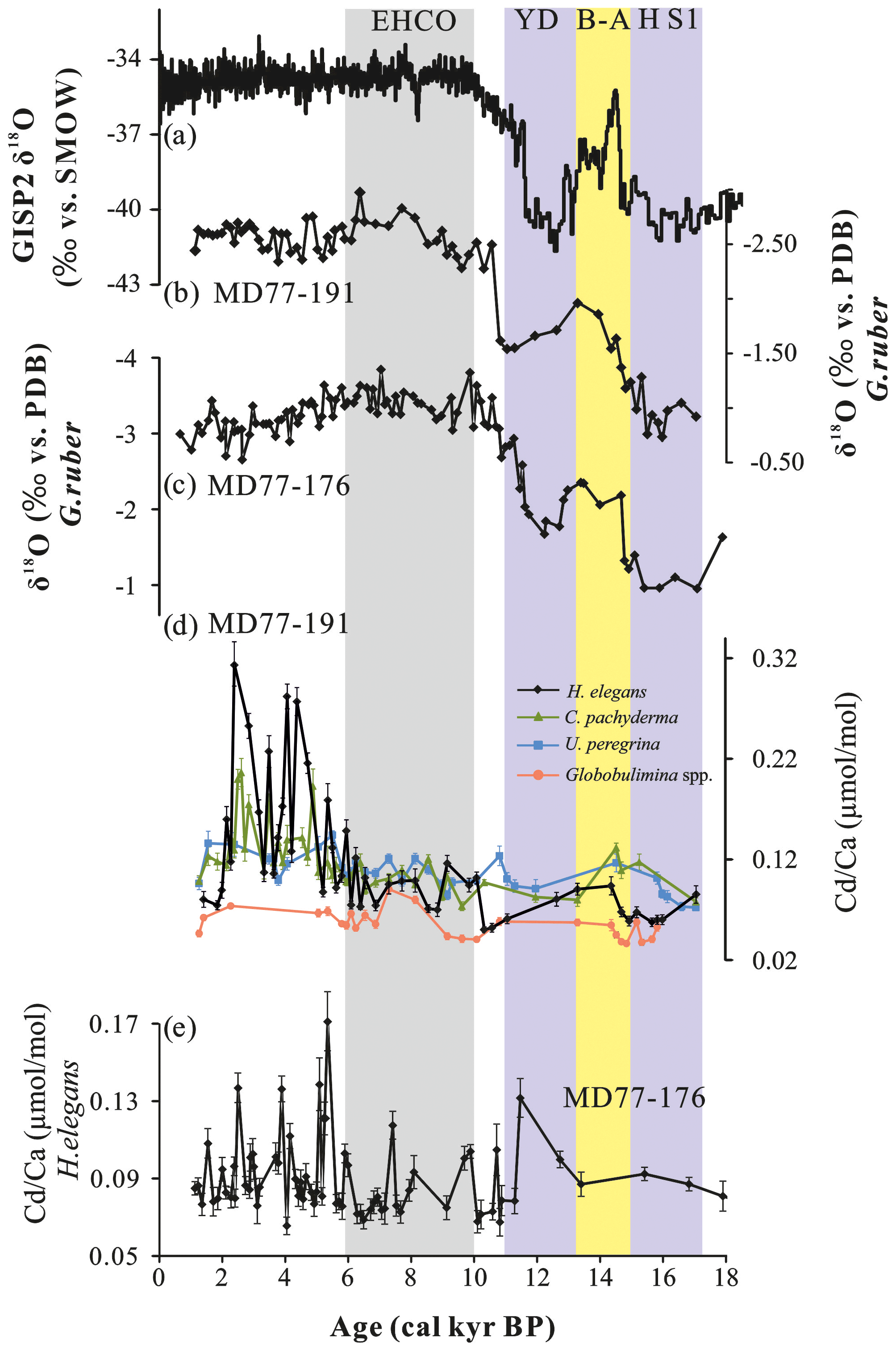
Figure 2(a) GISP2 Greenland ice core δ18O signal (Stuiver and Grootes, 2000). (b, c) Globigerinoides ruber δ18O records of cores MD77-191and MD77-176, respectively (Marzin et al., 2013; Ma et al., 2020). (d) records of the benthic foraminifera Hoeglundina elegans (black), Cibicidoides pachyderma (green), Uvigerina peregrina (blue) and Globobulimina spp. (orange) obtained from core MD77-191. (e) records of the benthic foraminifera H. elegans from core MD77-176. EHCO stands for Early Holocene Climate Optimum, YD for Younger Dryas, B-A for Bølling–Allerød and HS1 for Heinrich stadial 1.
The records for the calcite benthic species C. pachyderma and U. peregrina have very low time resolutions during the last deglaciation. However, some common patterns can be observed. The records of C. pachyderma and U. peregrina show lower values during the Heinrich stadial 1 (HS1, 17–15.2 cal ka) and the Younger Dryas (YD, 13–11 cal ka) cold periods, with average values of ∼0.08 µmol mol−1 for C. pachyderma and ∼0.09 µmol mol−1 for U. peregrina. By contrast, these two species display higher ratios (∼0.12 µmol mol−1) during the Bølling–Allerød warm period (B-A, 15–13.3 cal ka) compared with the HS1 and YD. Then, lower values (∼0.1 µmol mol−1 for C. pachyderma; 0.11 µmol mol−1 for U. peregrina) are observed during the early Holocene (10–5 cal ka) compared to larger variations occurring in the late Holocene (5.2–2.4 cal ka). The record of deep infaunal Globobulimina spp., obtained at a lower time resolution, shows different variations compared with the two other taxa without any clear trend during the Holocene.
The H. elegans values of core MD77-191 range from 0.05 to 0.31 µmol mol−1 since 17 cal kyr BP (Fig. 2d; Table S1). Depleted values at about 0.07 µmol mol−1 are recorded from the last deglaciation to the early Holocene (17–5 cal ka time interval). During the HS1 and the YD time intervals, a significant decrease of ∼0.06 µmol mol−1 occurred (even when taking into consideration the analytical error bar of ±0.02, 2σ), and a slight increase (0.09 µmol mol−1) is observed between 15 and 13.3 cal ka (B-A period). A rapid increase in the values beginning at 5.2 cal kyr BP reaches a maximum (0.31 µmol mol−1) during the late Holocene.
For core MD77-176, the H. elegans records range between 0.06 and 0.17 µmol mol−1 over the past 18 cal kyr BP (Fig. 2e; Table S1), without no clear trends and average benthic values of ∼0.09 µmol mol−1 during the different periods (HS1, YD and Holocene). However, the benthic record during the Holocene seems to exhibit a slight increase both in value and range of variations after 6 cal kyr BP.
4.2 Foraminifera assemblages of core MD77-191
Benthic foraminiferal species richness ranges between 16 and 36, and the total abundance fluctuates between 82 and 642 specimens (Table S2). Hyaline species are the dominant constituents (>80 %) and mainly consist of Bulimina aculeata, H. elegans, C. pachyderma, Uvigerina spp., Gyroidina broeckhiana, Globocassidulina subglobosa, Sphaeroidina bulloides, Gyroidinoides spp., Lenticulina spp., Melonis barleeanum and Globobulimina spp. (including Praeglobobulimina spp.) (in decreasing order of relative average abundance). Agglutinated taxa reach on average about 1.6 % and consist of Textularia sp., Martinottiella communis and Eggerella bradyi. The average percentage of porcelaneous species, characterized by Pyrgo elongata, Pyrgo murrhina, Pyrgo depressa, Pyrgoella irregularis, Quinqueloculina spp., Sigmoilopsis schlumbergeri and Spiroloculina spp., is about 5.1 %.
Furthermore, we merged species that share an ecological similarity, such as Globobulimina affinis, Globobulimina pacifica and Praeglobobulimina spp. into Globobulimina spp. A total of 74 samples and 55 groups and/or species were adopted to perform principal component analysis (PCA) in order to identify major faunal trends. The PCA analysis suggests that the benthic foraminifera could be grouped into three assemblages, with PC1 (positive and negative loadings) and PC2 (positive loadings) representing 42 % and 19 % of the total variance, respectively (Table 1). Moreover, compared with the total variance of PC1 and PC2, PC3 is the largest one and only explains 8 % of the total variance for the rest of the principal component scores (PCs). The species composition consists of H. elegans, Globobulimina spp. (positive loadings), Uvigerina peregrina and C. pachyderma (negative loadings) (Table 1). It seems that the main composition of assemblages (PC3) is quite similar to PC1 and does not show more information about the bottom conditions. Therefore, we only focus on PC1 and PC2 in the paper for the interpretation and do not present other PCs in the discussion.
Table 1Species composition of benthic foraminiferal assemblages from core MD77-191.

Assemblage 1 can be defined as the combination of Bulimina aculeata and C. pachyderma, together with Pullenia bulloides and Ehrenbergina trigona (Figs. 3 and S2) and display high positive PC1 loadings. This assemblage dominated the foraminiferal record during the late Holocene (between 6 and 1.4 cal ka).
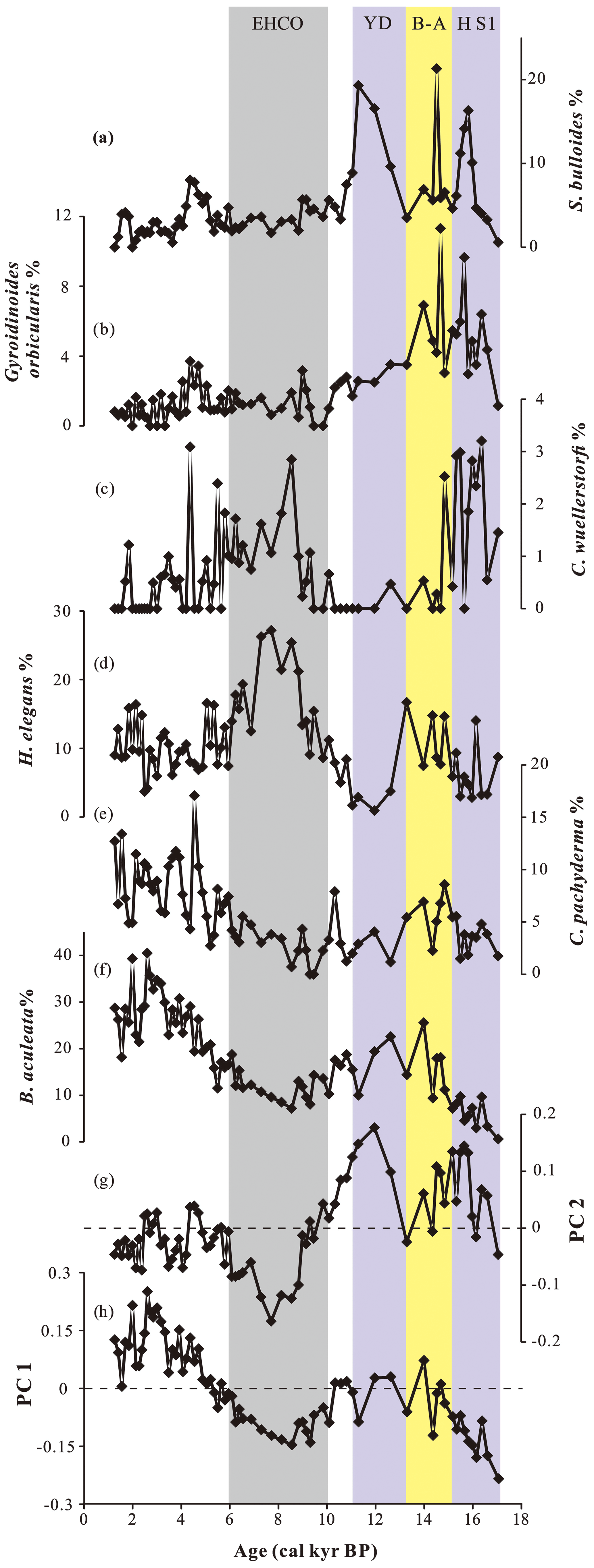
Figure 3Downcore variations in PC scores and the percentages of major species. (a) Sphaeroidina bulloides and (b) Gyroidinoides orbicularis are dominated assemblage 3, (c) Cibicidoides wuellerstorfi and (d) Hoeglundina elegans are the main associated species of assemblage 2, (e) Cibicidoides pachyderma and (f) Bulimina aculeata are major species in assemblage 1. The color-shaded intervals and abbreviations are the same as in Fig. 2.
By contrast, assemblage 2, dominated by H. elegans and Bulimina manginata, exhibits high negative PC1 loadings, and corresponds to the record during the early Holocene (Figs. 3 and S2). Other quantitatively important contributors are C. wuellerstorfi and Globocassidulina subglobosa (Fig. S2).
Then, assemblage 3, dominated by Sphaeroidina bulloides and Gyroidinoides orbicularis, corresponds to the positive loadings of PC2, which is more important during the last deglaciation (Figs. 3 and S2). The associated species of assemblage 3 are Bulimina mexicana and Gyroidinoides soldanii (Fig. S2).
However, as the main composition of PC2 negative loadings is dominated by the same benthic species in assemblages 1 and 2, it is difficult to glean any additional information from this analysis. Thus, to clarify the discussion, we prefer to use three assemblages in the following rather than the two PCs.
5.1 Past intermediate-water Cdw concentrations from the northern Indian Ocean
In the modern ocean, benthic foraminifera shows a positive correlation with Cdw and dissolved nutrients (phosphate and nitrate) (Boyle et al., 1976; Hester and Boyle, 1982). As aragonitic benthic foraminifera H. elegans faithfully records the bottom-water Cd concentrations (Cdw), ratios can be converted to seawater Cdw with the appropriate relationship (Eq. 1), where the partition coefficient Dp≈1 for all water depths (Boyle et al., 1995; Bryan and Marchitto, 2010).
In contrast, the partition coefficient for calcite species changes with water depth. For water depths between 1150 and 3000 m, Dp was calculated based on the equation of Boyle (1992; Eq. 2). The seawater Ca concentration is assumed to be at a constant mean value of 0.01 mol kg−1 (Boyle, 1992).
The intermediate Cdw results based on the H. elegans values of core MD77-191 range from 0.5 to 3.1 nmol kg−1 since 17 cal kyr BP (Fig. 4a), with a core top value of 0.80 nmol kg−1 in agreement with the estimated intermediate water depth modern Cdw (∼0.83 nmol kg−1) in the northern Indian Ocean (Boyle et al., 1995). The intermediate Cdw was also calculated from calcite benthic species C. pachyderma, U. peregrina and Globobulimina spp. from core MD77-191, with values ranging between 0.53–1.48, 0.52–1.04 and 0.26–0.65 µmol mol−1, respectively (Fig. 4a). The Cdw values of C. pachyderma and U. peregrina are within the same range. However, the H. elegans Cdw values are higher than those from the two calcite species, especially during the late Holocene. Moreover, the core top data of C. pachyderma and U. peregrina are also lower (∼0.7 and 0.69 nmol kg−1, respectively) than the modern estimated Cdw data (∼0.83 nmol kg−1) in the northern Indian Ocean (Boyle et al., 1995) (Fig. 4a). These depleted Cdw values may be related to the benthic foraminiferal microhabitat effect; indeed, U. peregrina is known to be strictly a shallow infaunal species, as well as C. pachyderma (Fontanier et al., 2002), differing from strictly epifaunal taxa, such as Cibicidoides wuellerstorfi (Mackensen et al., 1993).
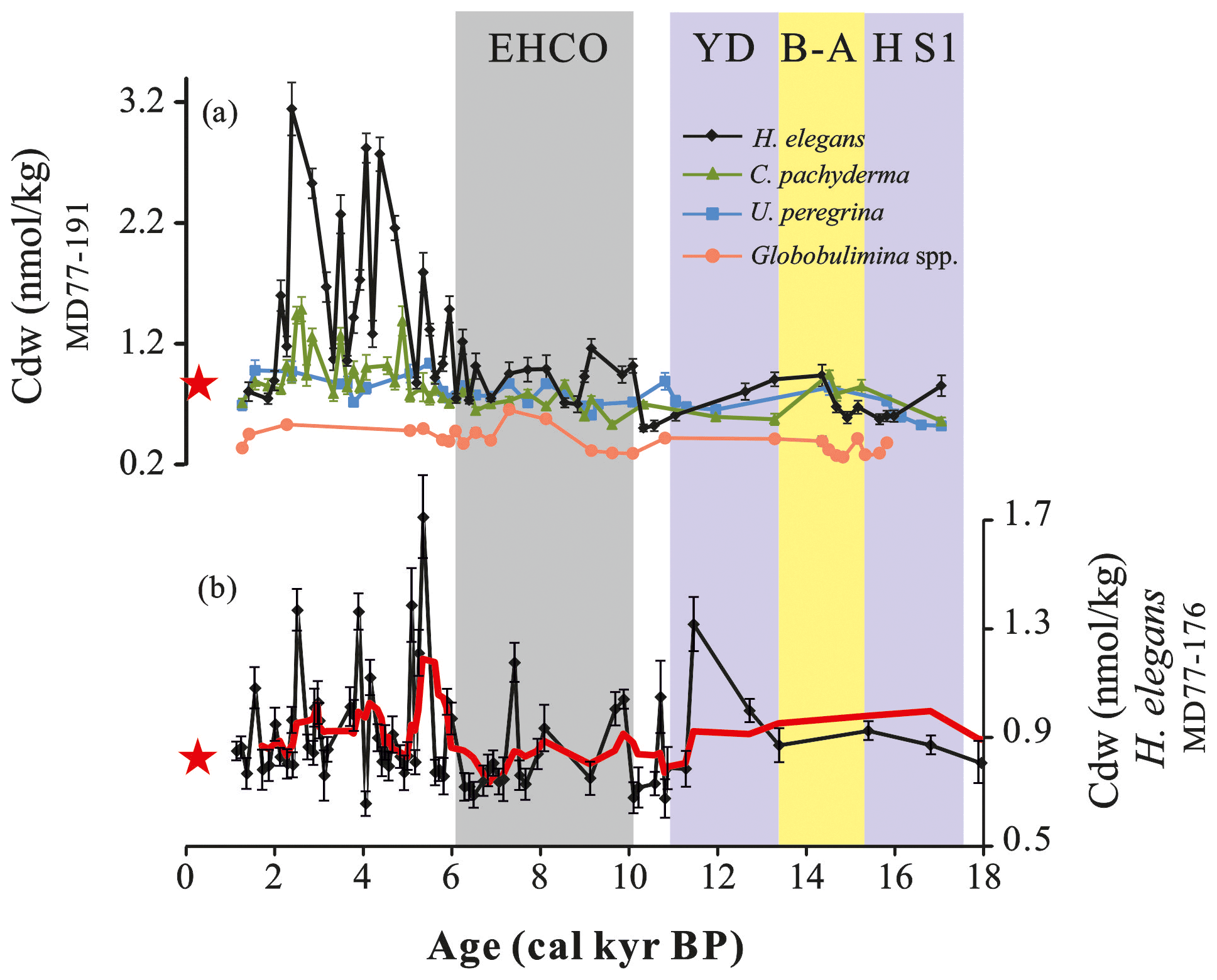
Figure 4(a) Cdw records calculated based on the of benthic foraminifera Hoeglundina elegans (black), Cibicidoides pachyderma (green), Uvigerina peregrina (blue) and Globobulimina spp. (orange) obtained from core MD77-191. (b) Cdw record from core MD77-176 reconstructed using H. elegans ; the red line is the smoothed curves using a two-point moving average. The red stars represent the modern Cdw (∼0.83 nmol kg−1) in the northern Indian Ocean (Boyle et al., 1995). The color-shaded intervals and abbreviations are the same as in Fig. 2.
Moreover, the deep infaunal Globobulimina spp. Cdw displays relatively much lower values and does not exhibit strong variations compared to the other species investigated in this study, displaying a general increasing trend from the last deglaciation to the Holocene. As Globobulimina spp. correspond to deep benthic infaunal species, this result may indicate a stable nutrient content of pore water as compared to other benthic taxa associated with bottom water (Fig. 4a). Thus, when tracking past changes in the bottom-water Cdw concentrations, the use of a strictly epifaunal species living at the water–sediment interface such as H. elegans appears to be more robust than using endofaunal species that live in contact with pore water.
Relative variations in the Cdw obtained from C. pachyderma and U. peregrina are in good agreement with the records obtained on H. elegans. Variations in H. elegans Cdw during the last deglaciation indicate a decrease of about ∼0.6 nmol kg−1 in the HS1 and YD periods, with a slight increase (0.9 nmol kg−1) during the warm B-A. Cdw results from core MD77-191 indicate a shift from the last deglaciation (∼0.7 nmol kg−1) to the late Holocene (∼1.59 nmol kg−1). During the Holocene, the Cdw records display relatively low values of around 0.9 nmol kg−1 in the 10–6 cal ka time interval and show a major shift at around 6.4 cal kyr BP with values rising up to 3.1 nmol kg−1.
For core MD77-176, the intermediate-water Cdw calculated from the H. elegans records ranges between 0.6 and 1.7 nmol kg−1 over the past 18 cal kyr BP (Fig. 4b). Compared with intermediate Cdw from MD77-191, the Cdw record of core MD77-176 does not display any clear trend from the last deglaciation to the Holocene. However, a slight increase is observed since 6 cal kyr BP, in agreement with the MD77-191 intermediate Cdw records. In addition, even though the MD77-176 record has a lower time resolution, it displays a shorter maximum (1.3 nmol kg−1) during the 13.4–11 cal ka time interval.
To summarize, among the three calcite benthic taxa and the aragonitic benthic species H. elegans, the records of H. elegans appear to be the most suitable for tracking past Cdw changes at intermediate water depth through time. Thus, in the following discussion, we will only focus on the intermediate Cdw calculated from the H. elegans from both studied cores.
5.2 Comparison between geochemical records and benthic foraminiferal assemblages
Comparing the geochemical records to the benthic assemblages, we can observe similar patterns. For core MD77-191 from the southeastern Arabian Sea, three benthic assemblages were identified since the last deglaciation. S. bulloides and Gyroidinoides orbicularis are major components of assemblage 3 (during the last deglaciation), together with B. mexicana and Gyroidinoides soldanii (Figs. 3 and S2). S. bulloides and B. mexicana are found in intermediate to high organic carbon flux rate regions (e.g., Schmiedl et al., 2000; Eberwein and Mackensen, 2006, 2008), while G. orbicularis and G. soldanii are associated with well-oxygenated and oligotrophic environments (Peterson, 1984; Burmistrova and Belyaeva, 2006; De and Gupta, 2010). Thus, assemblage 3 reflects mesotrophic environments and/or well-ventilated conditions during the last deglaciation. Although millennial-scale changes in the benthic foraminiferal assemblages during the last deglaciation could not be observed, assemblage 3 seems at least partly consistent with previous studies in the northern Indian Ocean based on multiple geochemical proxies (e.g., benthic δ13C, intermediate water [CO] and εNd records); these studies have revealed the presence of better-ventilated waters, which might correspond to AAIW, during the HS1 and YD (e.g., Yu et al., 2018; Ma et al., 2019, 2020).
Benthic foraminiferal assemblage 2 predominates during the early Holocene and is characterized by H. elegans and B. manginata as major contributors (Figs. 3 and S2). The other important contributors are C. wuellerstorfi and G. subglobosa. B. manginata is found in high organic carbon flux rate conditions (De Rijk et al., 2000; Eberwein and Mackensen, 2006, 2008). However, previous studies on H. elegans, C. wuellerstorfi and G. subglobosa indicate that these species correspond to high levels of dissolved oxygen and oligotrophic settings (e.g., Altenbach et al., 1999; Fontanier et al., 2002; Murgese and De Deckker, 2005, 2007; De and Gupta, 2010). Periods dominated by these taxa probably indicate high oxygen levels and an oligotrophic environment. This is consistent with previous studies in the area, based on benthic foraminiferal δ13C and Δ14C age difference (e.g., Naqvi et al., 1994; Bryan et al., 2010) (Fig. S3). Indeed, the glacial to Holocene benthic δ13C shifts (0.35 ‰–0.4 ‰, vs. Pee Dee Belemnite, PDB) at intermediate–deep water depth in the northern Indian Ocean are interpreted as reflecting an increased contribution of better-ventilated deep water NADW in IDW, during the Holocene (e.g., Naqvi et al., 1994; Ma et al., 2019) (Fig. S3). Furthermore, the increased benthic–planktonic (B-P) age offsets and depleted εNd records obtained from the same core site could also reflect the enhanced influence of NADW in IDW during the Holocene, which is characterized by well-ventilated conditions and depleted nutrient concentrations (modern Cdw, ∼0.2 nmol kg−1) (Poggemann et al., 2017; Yu et al., 2018; Ma et al., 2019). The impact of this change in the IDW composition can be recorded at intermediate water depth since the deep water masses are transformed to an upward flow during their pathway, thus being a potential contribution to intermediate-depth water masses (Naqvi et al., 1994; You, 2000). Although the intermediate benthic δ13C record from core MD77-191 is missing for the Last Glacial Maximum (LGM), the average value for the Holocene (∼0.31 ‰, vs. PDB) is consistent with previous studies carried out in the northern Indian Ocean; combined with the opposite trend between δ18Oivc records and intermediate-water temperature from MD77-191 (Ma et al., 2020), all these records suggest well-ventilated conditions (Fig. S3). To summarize, the predominance of benthic foraminifera assemblage 2 in the early Holocene seems to reflect better-ventilated water masses, related to an enhanced contribution of NADW in IDW at the core site, as already observed in previous studies (Poggemann et al., 2017; Yu et al., 2018; Ma et al., 2019, 2020).
By contrast, B. aculeata and C. pachyderma are major components of assemblage 1 (during the late Holocene), together with P. bulloides and E. trigona (Figs. 3 and S2). Living B. aculeata have a widespread distribution, with a preference for water depths ranging from 1500 to 2500 m, and are typically associated with high organic carbon fluxes (Mackensen et al., 1995; Almogi-Labin et al., 2000; Caulle et al., 2015). P. bulloides is a shallow infaunal species, which prefers mesotrophic environments and shows adaptability with respect to oxygen concentration in the Arabian Sea (Gupta and Thomas, 1999; Caulle et al., 2015). E. trigona is commonly recorded in low-oxygen habitats (Caulle et al., 2015). We thus interpret assemblage 1 as indicating relatively low-oxygen and meso- to eutrophic bottom-water conditions during the late Holocene (6–1.4 cal ka). However, the lower oxygen concentrations reflected by benthic fauna 1 seem to be the opposite of what would be expected under an enhanced influence of better-ventilated NADW in IDW during the Holocene in the northern Indian Ocean. Thus, another process has to be explored to combine our observations. To do that, we can use the relative abundance of Globigerina bulloides, a proxy for upwelling activity, that increased in the late Holocene in core MD77-191, suggesting an increased productivity in the southeastern Arabian Sea (Bassinot et al., 2011) (Fig. 5). This record is synchronous with the benthic foraminiferal assemblage 1 (during the late Holocene). Thus, increased surface productivity during the late Holocene could have introduced more organic matter into the intermediate water, leading to depleted oxygen conditions. By contrast, benthic assemblages 2 and 3 (during the last deglaciation and early Holocene; 17–6 cal ka) are associated with low G. bulloides abundances, suggesting lower productivity in the southeastern Arabian Sea during this period (Bassinot et al., 2011) and thus indicating that intermediate water masses were characterized by higher bottom-water oxygen conditions and a lower flux of organic matter. Therefore, all of these elements suggest that changes in primary productivity seem to be an important factor impacting the distribution of benthic assemblages at the core MD77-191 site, especially during the Holocene.
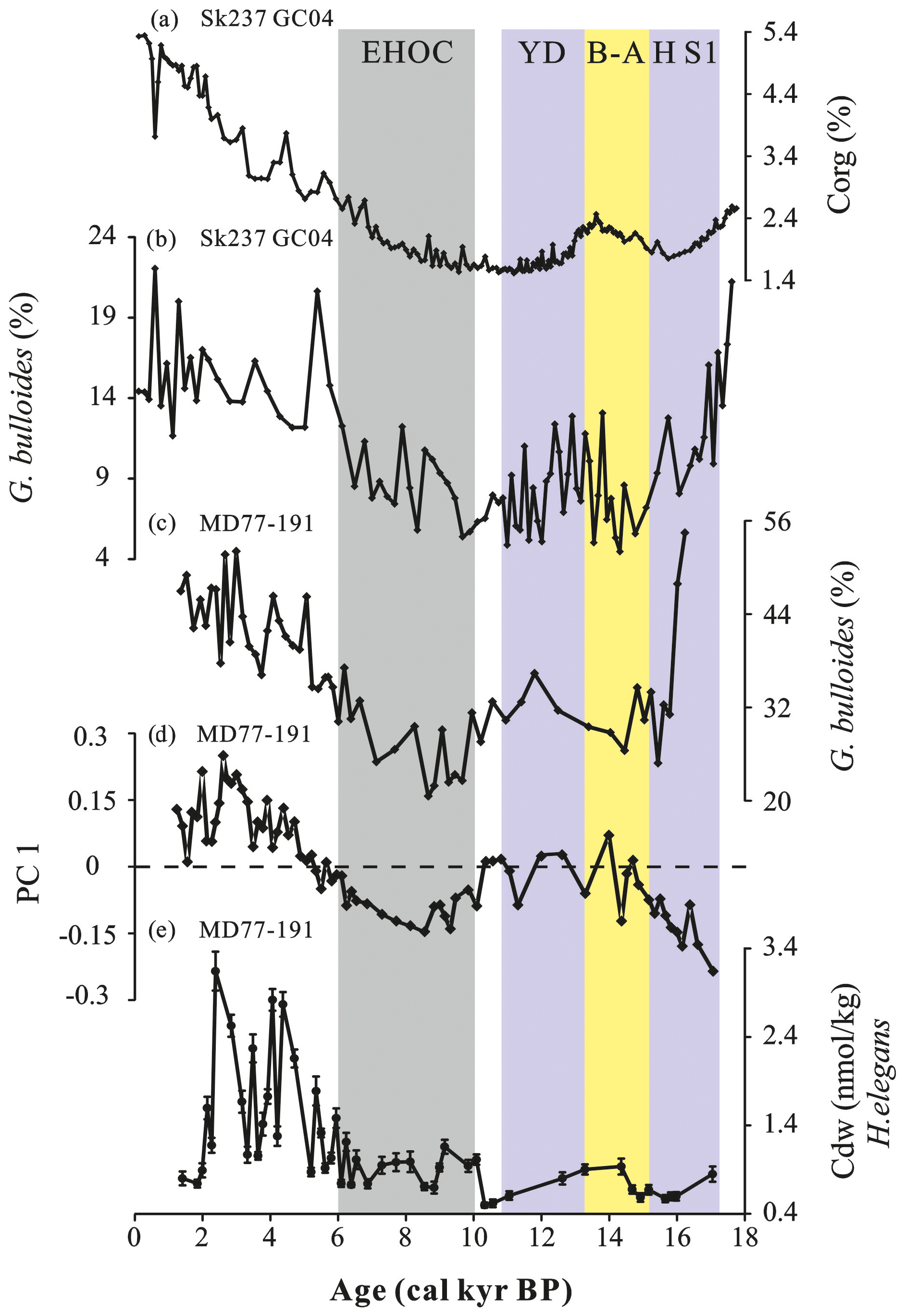
Figure 5(a) Organic carbon weight percentage (%Corg) and (b) G. bulloides percentage from core SK237 GC04 (1245 m, Arabian Sea, Naik et al., 2017). (c) Relative abundance of G. bulloides (Mléneck, 1997; Bassinot et al., 2011), (d) PC 1 scores and (e) Cdw records from core MD77-191 (Arabian Sea). The color-shaded intervals and abbreviations are the same as in Fig. 2.
In order to examine the relationships between intermediate Cdw and these different processes (surface productivity and/or water mass ventilation) in the eastern Arabian Sea, we can compare the MD77-191 Cdw values with the relative abundance of G. bulloides and benthic foraminiferal assemblage analyses from the same core MD77-191, together with the records for Corg and the G. bulloides percentage obtained from core SK237 GC04 (1245 m, southeastern Arabian Sea, Naik et al., 2017) (Fig. 5). Indeed, the total organic carbon (Corg) could also be used as a qualitative indicator of past productivity and/or bottom-water ventilation changes (Naidu et al., 1992; Canfield, 1994; Calvert et al., 1995; Naik et al., 2017). Despite a lower resolution for MD77-191 H. elegans Cdw records, when compared to the Corg and the G. bulloides percentage from core SK237 GC04, all of them seem to exhibit similar trends at a long timescale even though some small-scale discrepancies can be observed at a millennial timescales (Fig. 5). From the last deglaciation to the late Holocene, the Cdw record displays a significant shift from ∼0.7 nmol kg−1 to about twice the values of ∼1.59 nmol kg−1. The intermediate Cdw values are thus extremely high during the late Holocene and synchronous with the higher values of Corg and G. bulloides percentage records. These observed similar trends suggest that the increased surface productivity at the core site during the late Holocene is associated with higher intermediate Cdw values. Moreover, previous studies have suggested that increased Cdw values (>1 nmol kg−1) could correspond to elevated surface productivity (Bostock et al., 2010; Olsen et al., 2016). However, at a millennial timescale, we also observed several decreases in intermediate Cdw values (∼0.81 nmol kg−1) during the late Holocene, reaching nearly similar values during the last deglaciation (Fig. 5). Thus, the variations in the Cdw values cannot be fully associated with variations in the surface productivity.
As mentioned before, during the Holocene, an increased influence of NADW in IDW was observed in the northern Indian Ocean (Yu et al., 2018; Ma et al., 2019, 2020). NADW is characterized by a depleted nutrient content (modern Cdw, ∼0.2 nmol kg−1; Poggemann et al., 2017), and its contribution to IDW may affect the intermediate Cdw by deep water masses upwelling when flowing northward. However, during the late Holocene, benthic foraminiferal assemblage 1 is associated with lower oxygen concentrations, which seem to be inconsistent with an enhanced influence of better-ventilated NADW in IDW in the northern Indian Ocean. Therefore, this apparent discrepancy seems to indicate that deep–intermediate water mass variations are not an important control during the Holocene in this area, although we could not fully exclude the influence of NADW in IDW at a millennial timescale. Moreover, there is no clear evidence for such a millennial-scale variability in the IDW and/or NADW circulation in the studied area. Thus, we suggest the intermediate Cdw at the core MD77-191 site may be mainly influenced by surface productivity, especially during the Holocene.
In the Bay of Bengal, the benthic assemblages of core MD77-176 suggest that the intermediate water masses were characterized by oligotrophic to mesotrophic conditions and/or well-ventilated environments during the Holocene (Ma et al., 2019), associated with much lower surface productivity (Fig. S4). This observation is in agreement with low primary productivity during the Holocene reconstructed by the relative abundance of coccolith species Florisphaera profunda from the same core MD77-176 in the northeastern BoB (Zhou et al., 2020). In the modern ocean, Prasanna Kumar et al. (2001) indicate that primary productivity in the BoB is much lower than in the Arabian Sea, the lower surface productivity resulting from the large freshwater input from rivers and direct rainfall resulting from enhanced Indian summer monsoon precipitation (e.g., Vinayachandran et al., 2002; Madhupratap et al., 2003; Gauns et al., 2005). Moreover, when we compare the average Cdw value of core MD77-176 from the BoB (∼0.9 nmol kg−1) with results from core MD77-191 in the Arabian Sea (∼1.2 nmol kg−1), lower values, especially during the late Holocene, are in agreement with the benthic assemblages.
To sum up, variations in the benthic assemblages seem to be associated with changes in the deep water mass ventilation and/or organic matter flux, linked to surface productivity. The benthic foraminiferal fauna are consistent with the Cdw record of core MD77-191 particularly during the late Holocene (6–1.4 cal ka). Thus, our results seem to show that the Cdw record is mainly controlled by changes occurring at the surface, especially during the Holocene. However, at millennial timescales, such during the HS1 and YD, the percentages of planktonic species G. bulloides from cores MD77-191 and SK237 GC04 all indicate modest paleo-productivity, the opposite of what is suggested by the results of core MD77-191 Cdw and the Corg record obtained from core SK237 GC04. This interval is also marked by enriched G. ruber δ18O values, indicating a weaker monsoon and reduced freshwater inputs (Naik et al., 2017). This apparent discrepancy may be related to changes in the intermediate water mass sources and/or ventilation during the last deglaciation.
So, in the next sections, we discuss (i) processes controlling surface productivity and (ii) changes in the intermediate-water circulation, both of them being potential drivers of the observed variations.
5.3 Relationships between primary productivity and monsoon intensity
During the Holocene, the intermediate-water Cdw records obtained from cores MD77-191 and MD77-176 seem to display depleted values in the early Holocene, followed by an abrupt increasing trend at the middle Holocene and then reaching higher values on average (despite a short-timescale variability) during the late Holocene.
Of the two cores, core MD77-176, located in the northeastern BoB, shows the lowest intermediate Cdw (down to ∼0.83 nmol kg−1) during the 10–6 cal ka time interval. Observations described above suggest that this low in Cdw resulted from low primary productivity and thus reduced fluxes of organic matter to the intermediate depths. We attribute this evolution to monsoon variation. Indeed, the early Holocene Climate Optimum (10–6 cal ka) is characterized by enhanced monsoon precipitation (Marzin et al., 2013; Contreras-Rosales et al., 2014) (Fig. 6d–f) that resulted in increased freshwater discharge from the Ganges–Brahmaputra river system and from the Irrawaddy River. However, the distribution of chlorophyll in surface water of the western BoB suggests a low annual productivity, indicating that the BoB is not significantly influenced by the riverine nutrient input (Zhou et al., 2020). Thus, it is likely that this increase in freshwater drove pronounced ocean stratification in the northeast BoB, which could impede the nutrient transfer from the intermediate and/or deep layer to the euphotic upper seawater column and then induce low productivity. A similar low in Cdw values is observed in the reconstructed intermediate-water Cdw record from core MD77-191 during the early Holocene, with values descending to ∼0.92 nmol kg−1 in the 10–6 cal ka time interval. These low values of intermediate Cdw are coeval with low surface productivity as recorded by the G. bulloides percentage and low values in Corg content from SK237 GC04 in the Arabian Sea (Fig. 5). These variations are also recorded in changes in benthic assemblages, with the occurrence of assemblage 2 associated with high oxygen levels and an oligotrophic environment (Fig. 3). Off the southern tip of India, we cannot reject the possibility that increased monsoon precipitation and enhanced freshwater runoffs in the BoB during the early Holocene, inducing a stronger stratification, could explain part of the decrease in surface primary productivity. Yet, at this site, another explanation prevails which is related to the decrease in summer monsoon wind intensity that drives local Ekman pumping. As shown by Bassinot et al. (2011), the productivity variations at the southern tip of India are inversely related to the evolution of upwelling activity along the Oman Margin, to the west of the Arabian Sea. Based on a data–model comparison, Bassinot et al. (2011) showed that this anti-correlation can be attributed to the northward shift in the intertropical convergence zone (ITCZ) when boreal summer insolation reached a maximum in the early Holocene (Fig. 6a); this ITCZ location results in enhanced summer monsoon wind intensity and an increase in the associated Ekman pumping in the west of the Arabian Sea and along the Oman margin, while it weakens at the southern tip of India. This process may thus induce a decrease in surface productivity in the southeastern Arabian Sea.
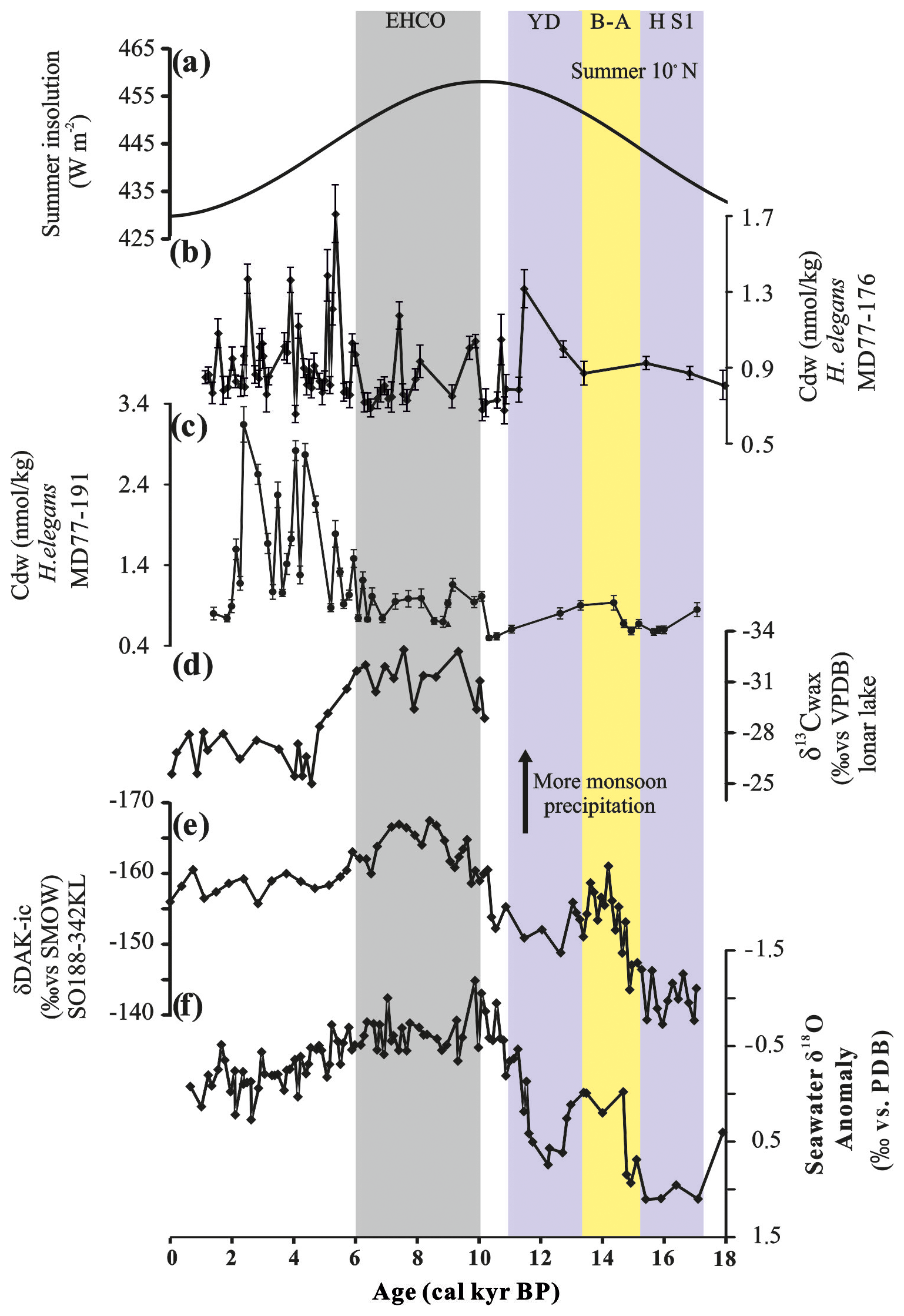
Figure 6(a) The solar insolation at 10∘ N in summer (Laskar et al., 2004). (b, c) Intermediate Cdw calculated from H. elegans obtained from MD77-176 and MD77-191, respectively. (d) Lonar Lake δ13Cwax record (Sarkar et al., 2015). (e) δDAlk−ic record from core SO188-342KL (Contreras-Rosales et al., 2014). (f) Seawater δ18O anomaly obtained from MD77-176 (Marzin et al., 2013). The color-shaded intervals and abbreviations are the same as in Fig. 2.
In addition, Naik et al. (2017) pointed out the co-existence of low productivity during the early Holocene in the BoB and to the south of India, in agreement with our data that clearly show the impact of such a reduction in surface primary productivity on the intermediate-water Cdw. These authors suggested a direct relationship between intense monsoon rainfall and reduced surface productivity. However, the northeastern BoB received a much larger amount of river input than the southern tip of India during the early Holocene (Marzin et al., 2013). Thus, it seems reasonable to propose that the northeastern BoB is more affected by the salinity-related stratification effect, while the southern tip of India is more affected by the decrease in wind intensity (Bassinot et al., 2011) with enhanced stratification being potentially made stronger by an additional freshwater effect, although weaker than in the BoB. Ultimately, both climatic features (summer wind intensity and precipitation) are directly under the control of monsoon evolution resulting from the orbital forcing of low-latitude boreal summer insolation.
By contrast, higher intermediate Cdw values from core MD77-191 associated with higher G. bulloides relative abundances and Corg from core SK237 GC04 during the 5.2–2.4 cal ka time interval could indicate enhanced productivity during the mid to late Holocene (Naik et al., 2017) (Fig. 5). To a lesser extent, this is also observed in the records from the northern BoB for the same time period. These changes are consistent with the weakened summer monsoon intensity, with less rainfall during the late Holocene, as observed in the BoB using core MD77-176 seawater δ18O and core SO188-342KL δDAlk−ic records (Marzin et al., 2013; Contreras-Rosales et al., 2014; Fig. 6e–f). In addition, this is also strongly supported by the δ13Cwax records from the Lonar Lake on the Indian continent (Sarkar et al., 2015; Fig. 6d) and a progressive increase in monsoon summer winds to the south of India (Bassinot et al., 2011). These observations could also strongly support the hypothesis that the major control on surface productivity is linked to monsoon evolution in the BoB and at the southern tip of the Arabian Sea during the Holocene (Bassinot et al., 2011; Naik et al., 2017; Zhou et al., 2020).
5.4 Millennial-scale changes in intermediate-water circulation during the deglaciation
During the last deglaciation, short events have been recorded at the site of core MD77-191 during the 16–15.2 (HS1) and 12.6–11 cal ka (YD) time intervals (Fig. 5). The low Cdw values in the MD77-191 record are coeval with reductions in Corg in core SK237 GC04 during the HS1 and YD periods (Fig. 5). According to previous studies, extremely high Cdw values (>1 nmol kg−1) were reported to have been associated with enhanced surface productivity (Bostock et al., 2010; Olsen et al., 2016). However, the range of values of intermediate Cdw (0.58–0.85 nmol kg−1, HS1; 0.5–0.8 nmol kg−1, YD) from core MD77-191 during the last deglaciation is much lower compared with the Holocene Cdw values (>1 nmol kg−1) and thus may be associated with other processes such as a better ventilation, changes in the water mass source and/or depleted surface productivity (Fig. 6). Significant decreases in G. bulloides relative abundance of SK237 GC04 (Naik et al., 2017) and MD77-191 records were observed from the HS1 to B-A (Bassinot et al., 2011), and thereafter slight increases occurred in the YD (Fig. 5). These high values at both core sites during the HS1 and YD may indicate an enhanced surface productivity during these intervals (Fig. 5). This should have led to increased intermediate Cdw and organic matter preservation under low oxygen concentration conditions during the HS1 and YD. However, despite a low resolution for the MD77-191 Cdw record during the last deglaciation, we do not observe high values of intermediate Cdw during the HS1 and YD (∼0.7 nmol kg−1) compared with the late Holocene (∼1.59 nmol kg−1), especially at 16.5–16 cal ka. Although we cannot fully discard the influence of surface productivity on the intermediate Cdw in these time intervals, this apparent discrepancy seems to provide another piece of evidence for the influence of changes in water masses and/or ventilation during the HS1 and YD, in line with previous studies and proxies in the northern Indian Ocean (Bryan et al., 2010; Yu et al., 2018; Ma et al., 2019, 2020).
Moreover, an increase in benthic δ13C values is observed during the HS1 and YD in the northern Indian Ocean (e.g., Duplessy et al., 1984; Curry et al., 1988; Naqvi et al., 1994; Jung et al., 2009; Ma et al., 2019) (Fig. S3). The increase in the different benthic δ13C records during the HS1 and YD in the western Arabian Sea, Pacific Ocean and BoB is interpreted as reflecting the northward expansion of AAIW (Pahnke and Zahn, 2005; Jung et al., 2009; Ma et al., 2019) (Fig. S3). The decreased B-P age obtained from marine sediment cores from the Arabian Sea and the Bay of Bengal during the same intervals could confirm enhanced vertical mixing in the Southern Ocean (Bryan et al., 2010; Ma et al., 2019). The transition in the εNd and Δ14C records during the deglaciation also indicates a strong northward penetration of AAIW within the North Atlantic and Bay of Bengal (e.g., Cao et al., 2007; Pahnke et al., 2008; Pena et al., 2013; Yu et al., 2018). In addition, during the HS1 and YD, a decrease in the [CO] record from core MD77-191 also suggests the release of CO2 from the deep ocean in the deglacial period through the expansion of AAIW (Ma et al., 2020). These time intervals are associated with better ventilation in the Southern Ocean (e.g., Anderson et al., 2009; Skinner et al., 2010), which led to enhanced vertical ventilation resulting in increased production of intermediate water masses (AAIW) (Anderson et al., 2009).
As mentioned before, previous studies have suggested an enhanced northward flow of southern sourced intermediate water mass AAIW, observed also in the Atlantic, Pacific and Indian oceans during the last deglaciation (e.g., Pahnke et al., 2008; Bryan et al., 2010; Poggemann et al., 2017; Yu et al., 2018; Ma et al., 2019, 2020), indicating that the source of intermediate water masses may be partly the same in these oceans. Thus, as the benthic δ13C values collected from the north Indian Ocean could better constrain the influence of AAIW in the two studied cores (Naqvi et al., 1994; Jung et al., 2009; Ma et al., 2019, 2020) (Fig. S3), we can also compare the range values of AAIW Cdw from both studied cores with data from the Atlantic and Pacific oceans at intermediate water depth during the HS1 and YD (Cdw, 0.3–0.9 nmol kg−1; Umling et al., 2018; Valley et al., 2017). Thereafter, we could get the ranges of Cdw–δ13C values of AAIW during these intervals, based on the benthic δ13C records in the Indian Ocean (Naqvi et al., 1994; Jung et al., 2009; Ma et al., 2019, 2020) as well as benthic Cdw values from the Pacific and Atlantic oceans (Valley et al., 2017; Umling et al., 2018) at intermediate water depths (Fig. 7). Unfortunately, the resolution of both intermediate Cdw and benthic δ13C from core MD77-176 (northeastern BoB) are very low for the HS1 and YD events, making it difficult to extract reliable information. Thus, we have decided to focus on the results from core MD77-191 (0.5–0.85 nmol kg−1) during these two time intervals; these results are in good agreement with the collected dataset (Fig. 7). Thus, the benthic Cdw results provide new evidence for tracking the northern flow of AAIW in the northern Indian Ocean, which increased during the HS1 and the YD.
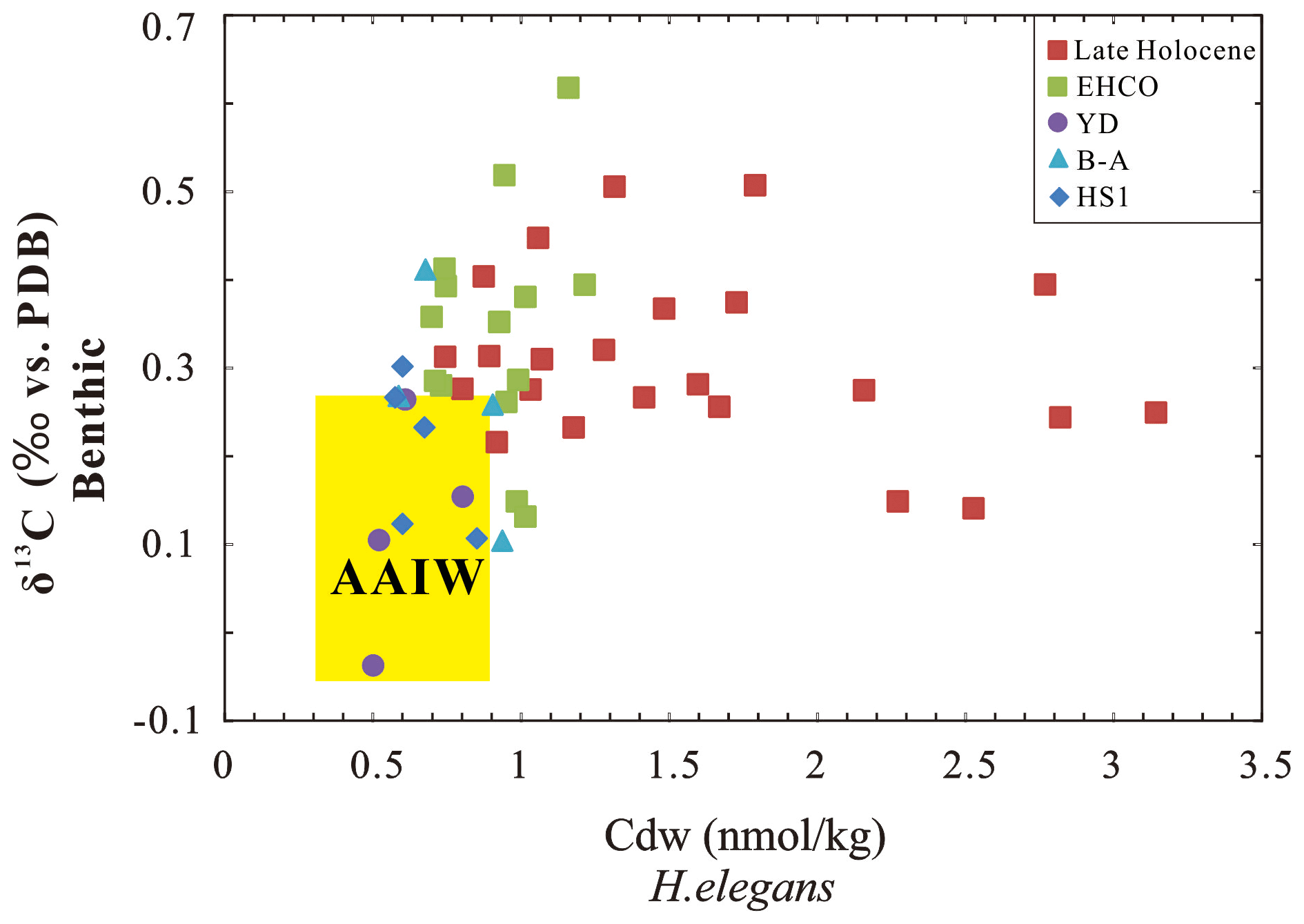
Figure 7Intermediate Cdw versus benthic δ13C obtained from core MD77-191 located off the southern tip of India. The yellow shaded area represents the ranges of Cdw–δ13C values of AAIW during the HS1 and YD, which were reconstructed in the Indian Ocean (benthic δ13C, Naqvi et al., 1994; Jung et al., 2009; Ma et al., 2019, 2020) and the Pacific and Atlantic oceans (benthic Cdw, Valley et al., 2017; Umling et al., 2018) at intermediate water depths. The abbreviations are the same as in Fig. 2.
Taken together, Cdw, the B-P age offset, benthic δ13C, εNd and Δ14C records reported from the northern Indian Ocean all suggest strong upwelling and enhanced northern flow of AAIW from the Southern Ocean during the HS1 and the YD. Thus, the variations in these records can provide strong evidence for the hypothesis that Southern Ocean upwelling played a vital role in the increase in atmospheric CO2 in the deglacial period (Anderson et al., 2009; Skinner et al., 2010, 2014). However, Kohfeld et al. (2005) suggested that although physical processes (such as ventilation) are involved in the glacial–interglacial atmospheric CO2 change, the biological pump may also contribute nearly half of the observed changes in CO2 during the glacial–interglacial transitions. As shown above, the HS1 event is characterized by reduced surface productivity, as revealed by the lower percentage values of G. bulloides in core MD77-191 (Bassinot et al., 2011) and by several studies of cores located in the eastern and western Arabian Sea within the Oxygen Minimum Zone (e.g., Schulz et al., 1998; Altabet et al., 2002; Ivanochko et al., 2005; Singh et al., 2006, 2011; Naik et al., 2017). This reduced productivity at a millennial timescale suggests that the entire biological factory was related to the reduced monsoon intensity during the North Atlantic Heinrich events (e.g., Singh et al., 2011; Naik et al., 2017). Thus, a weaker biological production could also have contributed to the two-step increase in atmospheric CO2 during the last deglaciation, at least for the HS1 period.
Changes in benthic foraminiferal and assemblages were reconstructed on core MD77-191 (1254 m water depth) located off the southern tip of India, as well as on core MD77-176 (1375 m water depth) from the northern BoB, in order to reveal the evolution of intermediate-water circulation and paleo-nutrient changes in the northern Indian Ocean since the last deglaciation. We reconstructed seawater Cdw concentration by converting H. elegans . Benthic ratios are mainly influenced by changes in surface productivity and intermediate–bottom-water ventilation.
Results indicate that assemblages 2 and 3, reflecting high bottom-water oxygen conditions and a low flux of organic matter, dominated between 17 and 6 cal ka, corresponding to a poor-productivity time period. The typical late Holocene assemblage indicates a relatively low oxygen level and meso- to eutrophic deep-water conditions, associated with high surface productivity. The early Holocene (10–6 cal ka) corresponds to a low in productivity associated with depleted Cdw in intermediate water. These observations seem to result from enhanced monsoon precipitation and increased river inputs from the Himalayan rivers, which led to more marked stratification in the BoB and a reduction in primary and export productivity. At the southern tip of India, the decrease in vertical mixing is also associated with a reduction in summer wind forcing resulting from the northward displacement of the ITCZ during summer (Bassinot et al., 2011). During the late Holocene (5.2–2.4 cal ka), the increased intermediate Cdw concentrations of cores MD77-191 and MD77-176 indicate enhanced surface productivity in the southeastern Arabian Sea and in the northeastern BoB, corresponding to weakened monsoon intensity and rainfall, in agreement with other local records and reconstructions of the paleo-monsoon strength. Thus, our results clearly show the strong control of intermediate-water Cdw during the Holocene by orbitally driven changes in summer monsoon productivity.
As far as millennial-scale variability is concerned, during the last deglaciation, decreased intermediate Cdw concentrations during the HS1 and the YD are coeval with increased benthic δ13C, depletion in [CO] and decreased B-P age offsets. These observations indicate that the low Cdw values in intermediate water mainly resulted from the increased northward flow of AAIW during the HS1 and YD intervals. These signals also provide strong evidence for the important role of enhanced Southern Ocean ventilation in the CO2 increase during the last deglaciation. The declined intermediate Cdw obtained from the southeastern Arabian Sea (Core MD77-191), combined with the published eastern and western Arabian Sea paleo-productivity results, provides evidence for the important influence of decreased monsoon intensity at a millennial timescale during cold events in the North Atlantic region, associated with the increase in atmospheric CO2 during the last deglaciation.
All data are given in Tables 1, S1 and S2 in the Supplement.
The supplement related to this article is available online at: https://doi.org/10.5194/cp-18-1757-2022-supplement.
RM, SS, FB and CC developed the idea and interpreted the results. CC and FB supplied foraminifera samples. RM did benthic foraminifera assemblage and geochemical analyses with the aide of FH and LL. ZY and LL joined the discussion. All co-authors helped to improve the article.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Ruifang Ma gratefully acknowledges the China Scholarship Council for providing funding for her study in France. The authors wish to thank one anonymous reviewer as well as André Bahr for useful suggestions and discussions during the revision process.
This research has been supported by the French National Research Agency under the “Investissements d'avenir” program (grant no. ANR-11-IDEX-0004-17-EURE-0006) and the CNRS (INSU-LEFE-IMAGO: Project CITRON GLACE).
This paper was edited by Arne Winguth and reviewed by André Bahr and one anonymous referee.
Almogi-Labin, A., Schmiedl, G., Hemleben, C., Siman-Tov, R., Segl, M., and Meischner, D.: The influence of the NE winter monsoon on productivity changes in the Gulf of Aden, NW Arabian Sea, during the last 530 ka as recorded by foraminifera, Mar. Micropaleontol., 40, 295–319, 2000.
Altabet, M. A., Higginson, M. J., and Murray, R. W.: The effect of millennial-scale changes in theArabian Sea denitrification on atmospheric CO2, Nature, 415, 159–162, 2002.
Altenbach, A. V., Pflaumann, U., Schiebel, R., Thies, A., Timm, S., and Trauth, M.: Scaling percentages and distributional patterns of benthic foraminifera with flux rates of organic carbon, J. Foramin. Res., 29, 173–185, 1999.
Anderson, R. F., Ali, S., Bradtmiller, L. I., Nielsen, S. H. H., Fleisher, M. Q., Anderson, B. E., and Burckle, L. H.: Wind-driven upwelling in the Southern Ocean and the deglacial rise in atmospheric CO2, Science, 323, 1443–1448, 2009.
Banse, K.: Seasonality of phytoplankton chlorophyll in the central and northern Arabian Sea, Deep-Sea Res., 34, 713–723, 1987.
Barker, S., Greaves, M., and Elderfield, H.: A study of cleaning procedures used for foraminiferal Mg/Ca paleothermometry, Geochem. Geophy. Geosy., 4, 1–20, 2003.
Bassinot, F. C., Marzin, C., Braconnot, P., Marti, O., Mathien-Blard, E., Lombard, F., and Bopp, L.: Holocene evolution of summer winds and marine productivity in the tropical Indian Ocean in response to insolation forcing: data-model comparison, Clim. Past, 7, 815–829, https://doi.org/10.5194/cp-7-815-2011, 2011.
Bauska, T. K., Baggenstos, D., Brook, E. J., Mix, A. C., Marcott, S. A., Petrenko, V. V., Schaefer, H., Severinghaus, J. P., and Lee, J. E.: Carbon isotopes characterize rapid changes in atmospheric carbon dioxide during the last deglaciation, P. Natl. Acad. Sci. USA, 113, 3465–3470, 2016.
Beal, L. M., Ffield, A., and Gordon, A. L.: Spreading of Red Sea overflow waters in the Indian Ocean, J. Geophys. Res.-Oceans, 105, 8549–8564, 2000.
Bostock, H. C., Opdyke, B. N., and Williams, M. J. M.: Characterising the intermediate depth waters of the Pacific Ocean using δ13C and other geochemical tracers, Deep-Sea Res. Pt. I, 57, 847–859, 2010.
Boyle, E. A.: Manganese carbonate overgrowths on foraminifera tests, Geochim. Cosmochim. Ac., 63, 353–353, 1983.
Boyle, E. A.: Cadmium: Chemical tracer of deepwater paleoceanography, Paleoceanography, 3, 471–489, 1988.
Boyle, E. A.: Cadmium and δ13C paleochemical ocean distributions during the stage 2 Glacial Maximum, Annu. Rev. Earth Pl. Sc., 20, 245–287, 1992.
Boyle, E. A. and Keigwin, L. D.: Deep circulation of the north Atlantic over the last 200,000 years: Geochemical evidence, Science, 218, 784–787, 1982.
Boyle, E. A., Sclater, F., and Edmond, J. M.: On the marine geochemistry of Cadmium, Nature, 263, 42–44, 1976.
Boyle, E. A., Labeyrie, L., and Duplessly, J. C.: Calcitic foraminiferal data confirmed by cadmium in aragonitic Hoeglundina: Application to the Last Glacial Maximum in the northern Indian Ocean, Paleoceanography, 10, 881–900, 1995.
Bryan, S. P. and Marchitto, T. M.: Testing the utility of paleonutrient proxies and in benthic foraminifera from thermocline waters, Geochem. Geophy. Geosy., 11, Q01005, https://doi.org/10.1029/2009GC002780, 2010.
Bryan, S. P., Marchitto, T. M., and Lehman, S. J.: The release of 14C-depleted carbon from the deep ocean during the last deglaciation: Evidence from the Arabian Sea, Earth Planet. Sc. Lett., 298, 244–254, 2010.
Burmistrova, I. I. and Belyaeva, N. V.: Bottom foraminiferal assemblages in the Deryugin Basin (Sea of Okhotsk) during the past 26000 years, Oceanology, 46, 834–840, 2006.
Calvert, S. E., Pedersen, T. F., Naidu, P. D., and von Stackelberg, U.: On the organic carbon maximum on the continental slope of the eastern Arabian Sea, J. Mar. Res., 53, 269–296, 1995.
Came, R. E., Oppo, D. W., Curry, W. B., and Lynch-Stieglitz, J.: Deglacial variability in the surface return flow of the Atlantic meridional overturning circulation, Paleoceanography, 23, PA1217, https://doi.org/10.1029/2007PA001450, 2008.
Canfield, D. E.: Factors influencing organic carbon preservation inmarine sediments, Chem. Geol., 114, 315–329, 1994.
Cao, L., Fairbanks, R. G., Mortlock, R. A., and Risk, M. J.: Radiocarbon reservoir age of high latitude north Atlantic surface water during the last deglacial, Quaternary Sci. Rev., 26, 732–742, 2007.
Caulle, C., Mojtahid, M., Gooday, A. J., Jorissen, F. J., and Kitazato, H.: Living (Rose-Bengal-stained) benthic foraminiferal faunas along a strong bottom-water oxygen gradient on the Indian margin (Arabian Sea), Biogeosciences, 12, 5005–5019, https://doi.org/10.5194/bg-12-5005-2015, 2015.
Contreras-Rosales, L. A., Jennerjahn, T., Tharammal, T., Meyer, V., Lückge, A., Paul, A., and Schefuß, E.: Evolution of the Indian Summer Monsoon and terrestrial vegetation in the Bengal region during the past 18 ka, Quaternary Sci. Rev., 102, 133–148, 2014.
Corliss, B. H.: Recent deep-sea benthonic foraminiferal distributions in the southeast Indian Ocean: Inferred bottom-water routes and ecological implications, Mar. Geol., 31, 115–138, 1979.
Corliss, B. H., Martinson, D. G., and Keffer, T.: Late Quaternary deep-ocean circulation, Geol. Soc. Am. Bull., 97, 1106–1121, 1986.
Curry, W. B., Duplessy, J. C., Labeyrie, L. D., and Shackleton, N. J.: Changes in the distribution of δ13C of deep water σCO2 between the last glaciation and the Holocene, Paleoceanography, 3, 317–341, 1988.
De, S. and Gupta, A. K.: Deep-sea faunal provinces and their inferred environments in the Indian Ocean based on distribution of recent benthic foraminifera, Palaeogeogr. Palaeocl., 291, 429–442, 2010.
Den Dulk, M., Reichart, G. J., Memon, G. M., Roelofs, E. M. P., Zachariasse, W. J., and Zwaan, G. J. V. D.: Benthic foraminiferal response to variations in surface water productivity and oxygenation in the northern Arabian Sea, Mar. Micropaleontol., 35, 43–66, 1998.
De Rijk, S., Jorissen, F. J., Rohling, E. J., and Troelstra, S. R.: Organic flux control on bathymetric zonation of Mediterranean benthic foraminifera, Mar. Micropaleontol., 40, 151–166, 2000.
Duplessy, J. C., Shackleton, N. J., Matthews, R. K., Prell, W., Ruddiman, W. F., Caralp, M., and Hendy, C. H.: 13C record of benthic foraminifera in the last interglacial ocean: Implications for the carbon cycle and the global deep water circulation, Quaternary Res., 21, 225–243, 1984.
Eberwein, A. and Mackensen, A.: Live and dead benthic foraminifera and test δ13C record primary productivity off Morocco (NW-Africa), Deep-Sea Res. Pt. I, 53, 1379–1405, 2006.
Eberwein, A. and Mackensen, A.: Last Glacial Maximum paleoproductivity and water masses off NW-Africa: Evidence from benthic foraminifera and stable isotopes, Mar. Micropaleontol., 67, 87–103, 2008.
Elderfield, H. and Rickaby, R. E. M.: Oceanic ratio and nutrient utilization in the glacial Southern Ocean, Nature, 405, 305–310, 2000.
Fontanier, C., Jorissen, F. J., Licari, L., Alexandre, A., Anschutz, P., and Carbonel, P.: Live benthic foraminiferal faunas from the Bay of Biscay: Faunal density, composition, and microhabitats, Deep-Sea Res. Pt. I, 49, 751–785, 2002.
Gauns, M., Madhupratap, M., Ramaiah, N., Jyothibabu, R., Fernandes, V., Paul, J. T., and Prasanna Kumar, S.: Comparative accounts of biological productivity characteristics and estimates of carbon fluxes in the Arabian Sea and the Bay of Bengal, Deep-Sea Res. Pt. II, 52, 2003–2017, 2005.
Gomes, H., Goes, J., and Saino, T.: Influence of physical processes and freshwater discharge on the seasonality of phytoplankton regime in the Bay of Bengal, Cont. Shelf Res., 20, 313–330, 2000.
Gupta, A. K. and Thomas, E.: Latest Miocene-Pleistocene productivity and deep-sea ventilation in the Northwestern Indian Ocean (Deep Sea Drilling Project Site 219), Paleoceanography, 14, 62–73, 1999.
Gupta, A. K., Anderson, D. M., and Overpeck, J. T.: Abrupt changes in the Asian Southwest Monsoon during the Holocene and their links to the North Atlantic Ocean, Nature, 421, 354–357, 2003.
Hammer, Ø., Harper, D. A. T., and Ryan, P. D.: Past: Paleontological statistics software package for education and data analysis, Palaeontol. Electron., 4, 9 pp., 2001.
Hermelin, J. O. R.: The benthic foraminiferal faunas of sites 725, 726, and 728 (Oman margin, northwestern Arabian Sea), in: Proceedings of the Ocean Drilling Program, Scientific Results, edited by: Prell, W. L., Niitsuma, N., Emeis, K.-C., Al-Sulaiman, Z. K., Al-Tobbah, A. N. K., Anderson, D. M., Barnes, R. O., Bilak, R. A., Bloemendal, J., Bray, C. J., Busch, W. H., Clemens, S. C., de Menocal, P., Debrabant, P., Hayashida, A., Hermelin, J. O. R., Jarrard, R. D., Krissek, L. A., Kroon, D., Murray, D. W., Nigrini, C. A., Pedersen, T. F., Ricken, W., Shimmield, G. B., Spaulding, S. A., Takayama, T., ten Haven, H. L., and Weedon, G. P., College Station, TX (Ocean Drilling Program), 117, 55–87, https://doi.org/10.2973/odp.proc.sr.117.130.1991, 1991.
Hermelin, J. O. R.: Variations in the benthic foraminiferal fauna of the Arabian Sea: A response to changes in upwelling intensity?, Geological Society, London, Special Publications, 64, 151–166, 1992.
Hermelin, J. O. R. and Shimmield, G. B.: Impact of productivity events on the benthic foraminiferal fauna in the Arabian Sea over the last 150,000 years, Paleoceanography, 10, 85–116, 1995.
Hertzberg, J. E., Lund, D. C., Schmittner, A., and Skrivanek, A. L.: Evidence for a biological pump driver of atmospheric CO2 rise during Heinrich Stadial 1, Geophys. Res. Lett., 43, 12242–12251, 2016.
Hester, K. and Boyle, E.: Water chemistry control of Cadmium content in recent benthic foraminifera, Nature, 298, 260–262, 1982.
Holbourn, A., Henderson, A. S., and Macleod, N.: Front matter, Atlas of benthic foraminifera, 1–641, https://doi.org/10.1002/9781118452493, 2013.
Ivanochko, T. S., Ganeshram, R. S., Brummer, G. J. A., Ganssen, G., Jung, S. J. A., Moreton, S. G., and Kroon, D.: Variations in tropical convection as an amplifier of global climate change at the millennial scale, Earth Planet. Sci. Lett., 235, 302–314, 2005.
Jaccard, S. L., Galbraith, E. D., Martínez-García, A., and Anderson, R. F.: Covariation of deep Southern Ocean oxygenation and atmospheric CO2 through the last ice age, Nature, 530, 207–210, 2016.
Jones, R. W.: The challenger foraminifera, Oxford University Press, https://doi.org/10.1046/j.1420-9101.1996.9010124.x, 1994.
Jung, S. J. A., Kroon, D., Ganssen, G., Peeters, F., and Ganeshram, R.: Enhanced Arabian Sea intermediate water flow during glacial North Atlantic cold phases, Earth Planet. Sc. Lett., 280, 220–228, 2009.
Kohfeld, K. E., Quéré, C. L., Harrison, S. P., and Anderson, R. F.: Role of marine biology in Glacial-interglacial CO2 cycles, Science, 308, 74–78, 2005.
Laskar, L., Robutel, P., Joutel, F., Gastineau, M., Correia, A. C., and Levrard, B.: A long-term numerical solution for the insolation quantities of the Earth, Astron. Astrophys., 428, 261–285, 2004.
Lévy, M., Shankar, D., André, J.-M., Shenoi, S., Durand, F., and De Boyer Montegut, C.: Basin-wide seasonal evolution of the Indian Ocean's phytoplankton blooms, J. Geophys. Res., 112, C12014, https://doi.org/10.1029/2007JC004090, 2007.
Loeblich, A. R. and Tappan, H.: Generic taxa erroneously regarded as foraminifers, in: Foraminiferal genera and their classification, edited by: Loeblich, A. R. and Tappan, H., Springer US, Boston, MA, 726–730, 1988.
Lynch-Stieglitz, J., Fairbanks, R. G., and Charles, C. D.: Glacial-interglacial history of Antarctic Intermediate Water: Relative strengths of Antarctic versus Indian Ocean sources, Paleoceanography, 9, 7–29, 1994.
Ma, R., Sépulcre, S., Licari, L., Bassinot, F., Liu, Z., Tisnérat-Laborde, N., Kallel, N., Yu, Z., and Colin, C.: Changes in intermediate circulation in the Bay of Bengal since the Last Glacial Maximum as inferred from benthic foraminifera assemblages and geochemical proxies, Geochem. Geophy. Geosy., 20, 1592–1608, 2019.
Ma, R., Sépulcre, S., Bassinot, F., Haurine, F., Tisnérat-Laborde, N., and Colin, C.: North Indian Ocean circulation since the last deglaciation as inferred from new elemental ratio records for benthic foraminifera Hoeglundina elegans, Paleoceanography and Paleoclimatology, 35, e2019PA003801, https://doi.org/10.1029/2019PA003801, 2020.
Mackensen, A., Hubberten, H. W., Bickert, T., Fischer, G., and Futterer, D. K.: δ13C in benthic foraminiferal tests of Fontbotia wuellerstorfi (Schwager) relative to δ13C of dissolved inorganic carbon in Southern Ocean deep water: implications for Glacial ocean circulation models, Paleoceanography, 6, 587–610, 1993.
Mackensen, A., Schmiedl, G., Harloff, J., and Giese, M.: Deep-sea foraminifera in the South Atlantic Ocean; ecology and assemblage generation, Micropaleontology, 41, 342–358, 1995.
Madhupratap, M., Gauns, M., Ramaiah, N., Prasanna Kumar, S., Muraleedharan, P. M., Sousa, S. N., and Muraleedharan, U.: Biogeochemistry of the Bay of Bengal: physical, chemical and primary productivity characteristics of the central and western Bay of Bengal during summer monsoon 2001, Deep-Sea Res. Pt. II, 50, 881–896, 2003.
Marchitto, T. M. and Broecker, W. S.: Deep water mass geometry in the glacial atlantic ocean: A review of constraints from the paleonutrient proxy , Geochem. Geophy. Geosy., 7, Q12003, https://doi.org/10.1029/2006GC001323, 2006.
Marchitto, T. M., Lehman, S. J., Ortiz, J. D., Flückiger, J., and Geen, A. V.: Marine radiocarbon evidence for the mechanism of deglacial atmospheric CO2 rise, Science, 316, 1456–1459, 2007.
Marra, J. and Barber, R. T.: Primary productivity in the Arabian Sea: A synthesis of JGOFS data, Prog. Oceanogr., 65, 159–175, 2005.
Marzin, C., Kallel, N., Kageyama, M., Duplessy, J.-C., and Braconnot, P.: Glacial fluctuations of the Indian monsoon and their relationship with North Atlantic climate: new data and modelling experiments, Clim. Past, 9, 2135–2151, https://doi.org/10.5194/cp-9-2135-2013, 2013.
Mléneck, V. M.: Sédimentation et dissolution des carbonates biogéniques aux moyennes latitudes Nord et Sud, Approche quantitative et relations avec les paléocirculations océaniques des derniers 150 000 ans, PhD thesis, Université Bordeaux I, 277 pp., 1997 (in French).
Monnin, E., Indermühle, A., Dällenbach, A., Flückiger, J., Stauffer, B., Stocker, T. F., Raynaud, D., and Barnola, J. M.: Atmospheric CO2 concentrations over the last glacial termination, Science, 291, 112–114, 2001.
Murgese, D. S. and De Deckker, P.: The distribution of deep-sea benthic foraminifera in core tops from the eastern Indian Ocean, Mar. Micropaleontol., 56, 25–49, 2005.
Murgese, D. S. and De Deckker, P.: The late quaternary evolution of water masses in the eastern Indian Ocean between Australia and Indonesia, based on benthic foraminifera faunal and carbon isotopes analyses, Palaeogeogr. Palaeocl., 247, 382–401, 2007.
Naidu, P. D. and Malmgren, B. A.: A high-resolution record of late Quaternary upwelling along the Oman margin, Arabian Sea based on planktonic foraminifera, Paleoceanography, 11, 129–140, 1996.
Naidu, P. D., Prakash Babu, C., and Rao, C. M.: The upwelling record in the sediments of the western continental margin of India, Deep-Sea Res., 39, 715–723, 1992.
Naik, D. K., Saraswat, R., Lea, D. W., Kurtarkar, S. R., and Mackensen, A.: Last glacial-interglacial productivity and associated changes in the eastern Arabian Sea, Palaeogeogr. Palaeocl., 483, 147–156, 2017.
Naqvi, W. A., Charles, C. D., and Fairbanks, R. G.: Carbon and oxygen isotopic records of benthic foraminifera from the northeast indian ocean: Implications on glacial-interglacial atmospheric CO2 changes, Earth Planet. Sc. Lett., 121, 99–110, 1994.
Olsen, A., Key, R. M., van Heuven, S., Lauvset, S. K., Velo, A., Lin, X., Schirnick, C., Kozyr, A., Tanhua, T., Hoppema, M., Jutterström, S., Steinfeldt, R., Jeansson, E., Ishii, M., Pérez, F. F., and Suzuki, T.: The Global Ocean Data Analysis Project version 2 (GLODAPv2) – an internally consistent data product for the world ocean, Earth Syst. Sci. Data, 8, 297–323, doi10.5194/essd-8-297-2016, 2016.
Olson, D. B., Hitchcock, G. L., Fine, R. A., and Warren, B. A.: Maintenance of the low-oxygen layer in the central Arabian Sea, Deep-Sea Res. Pt. II, 40, 673–685, 1993.
O'Malley, R.: Ocean productivity, College of Science, Oregon State University, USA, http://science.oregonstate.edu/ocean.productivity/index.php (last access: 1 December 2018), 2017.
Oppo, D. W. and Fairbanks, R. G.: Variability in the deep and intermediate water circulation of the Atlantic Ocean during the past 25,000 years: Northern Hemisphere modulation of the Southern Ocean, Earth Planet. Sc. Lett., 86, 1–15, 1987.
Pahnke, K. and Zahn, R.: Southern Hemisphere water mass conversion linked with north Atlantic climate variability, Science, 307, 1741–1746, 2005.
Pahnke, K., Goldstein, S. L., and Hemming, S. R.: Abrupt changes in Antarctic Intermediate Water circulation over the past 25,000 years, Nat. Geosci., 1, 870–874, 2008.
Pena, L. D., Goldstein, S. L., Hemming, S. R., Jones, K. M., Calvo, E., Pelejero, C., and Cacho, I.: Rapid changes in meridional advection of Southern Ocean intermediate waters to the tropical Pacific during the last 30 kyr, Earth Planet. Sc. Lett., 368, 20–32, 2013.
Peterson, L. C.: Recent abyssal benthic foraminiferal biofacies of the eastern Equatorial Indian Ocean, Mar. Micropaleontol., 8, 479–519, 1984.
Phillips, S. C., Johnson, J. E., Giosan, L., and Rose, K.: Monsoon-influenced variation in productivity and lithogenic sediment flux since 110 ka in the offshore Mahanadi Basin, northern Bay of Bengal, Mar. Petrol. Geol., 58, 502–525, 2014.
Pichevin, L. E., Reynolds, B. C., Ganeshram, R. S., Cacho, I., Pena, L., Keefe, K., and Ellam, R. M.: Enhanced carbon pump inferred from relaxation of nutrient limitation in the glacial ocean, Nature, 459, 1114–1117, 2009.
Poggemann, D. W., Hathorne, E., Nuernberg, D., Frank, M., Bruhn, I., Reißig, S., and Bahr, A.: Rapid deglacial injection of nutrients into the tropical Atlantic via Antarctic Intermediate Water, Earth Planet. Sc. Lett., 463, 118–126, 2017.
Prasanna Kumar, S., Madhupratap, M., Dileep Kumar, M., Muraleedharan, P. M., de Souza, S. N., Gauns, M., and Sarma, V. V. S. S.: High biological productivity in the central Arabian Sea during the summer monsoon driven by Ekman pumping and lateral advection, Curr. Sci. India, 81, 1633–1638, 2001.
Prell, W. L. and Kutzbach, J. L.: Monsoon variability over the past 150,000 years, J. Geophys. Res., 92, 8411–8425, 1987.
Reid, J. L.: On the total geostrophic circulation of the south Pacific Ocean: Flow patterns, tracers and transports, Prog. Oceanogr., 16, 1–61, 2003.
Saraswat, R., Nigam, R., and Correge, T.: A glimpse of the Quaternary monsoon history from India and adjoining seas, Palaeogeogr. Palaeocl., 397, 1–6, 2014.
Sarkar, S., Prasad, S., Wilkes, H., Riedel, N., Stebich, M., Basavaiah, N., and Sachse, D.: Monsoon source shifts during the drying mid-Holocene: Biomarker isotope based evidence from the core 'monsoon zone' (CMZ) of India, Quaternary Sci. Rev., 123, 144–157, 2015.
Schlitzer, R.: Electronic Atlas of WOCE Hydrographic and Tracer Data Now Available, Eos T. Am. Geophys. Un., 81, p. 45, https://doi.org/10.1029/00EO00028, 2000.
Schlitzer, R.: Ocean data view, ODV AWI, https://odv.awi.de/ (last access: 4 April 2022), 2015.
Schmiedl, G., Hemleben, C., Keller, J., and Segl, M.: Impact of climatic changes on the benthic foraminiferal fauna in the Ionian Sea during the last 330,000 years, Paleoceanography, 13, 447–458, 1998.
Schmiedl, G., De Bovee, F., Buscail, R., Charriere, B., Hemleben, C., Medernach, L., and Picon, P.: Trophic control of benthic foraminiferal abundance and microhabitat in the bathyal Gulf of Lions, western Mediterranean Sea, Mar. Micropaleontol., 40, 167–188, 2000.
Schnitker, D.: Deep-sea benthic foraminifers: Food and bottom water masses, in: Carbon cycling in the glacial ocean: Constraints on the ocean's role in global change, edited by: Zahn, R., Pedersent, T. F., Kaminski, M. A., and Labeyrie, L., NATO ASI Series (Series I: Global Environmental Change), vol. 17, Springer, Berlin, Heidelberg, https://doi.org/10.1007/978-3-642-78737-9_23, 1994.
Schott, F. A. and McCreary, J. P.: The monsoon circulation of the Indian Ocean, Prog. Oceanogr., 51, 1–123, 2001.
Schulz, H., von Rad, U., and Erlenkeuser, H.: Correlation between Arabian Sea and Greenland climate oscillation of the past 110,000 years, Nature, 393, 54–57, 1998.
Shankar, D., Vinayachandran, P. N., and Unnikrishnan, A. S.: The monsoon currents in the north Indian Ocean, Prog. Oceanogr., 52, 63–120, 2002.
Singh, A. D., Kroon, D., and Ganeshram, R.: Millennial scale variations in productivity and OMZ intensity in the eastern Arabian Sea, Journal of the Geological Society of India, 68, 369–377, 2006.
Singh, A. D., Jung, S. J. A., Darling, K., Ganeshram, R., Ivanochko, T., and Kroon, D.: Productivity collapses in the Arabian Sea during glacial cold phases, Paleoceanography, 26, PA3210, https://doi.org/10.1029/2009PA001923, 2011.
Skinner, L. C., Fallon, S., Waelbroeck, C., Michel, E., and Barker, S.: Ventilation of the deep Southern Ocean and deglacial CO2 rise, Science, 328, 1147–1151, 2010.
Skinner, L. C., Claire, W., Scrivner, A. E., and Fallon, S. J.: Radiocarbon evidence for alternating northern and southern sources of ventilation of the deep Atlantic carbon pool during the last deglaciation, P. Natl. Acas. Sci. USA, 111, 5480–5484, 2014.
Stuiver, M. and Grootes, P. M.: GISP2 oxygen isotope ratios, Quaternary Res., 53, 277–284, 2000.
Tachikawa, K. and Elderfield, H.: Microhabitat effects on and δ13C of benthic foramnifera, Earth Planet. Sc. Lett., 202, 607–624, 2002.
Talley, L. D., Pickard, G. L., Emery, W. J., and Swift, J. H.: Descriptive physical oceanography: An introduction, 6th edn., Academic Press, Boston, 1–383, https://doi.org/10.1016/C2009-0-24322-4, 2011.
Thushara, V. and Vinayachandran, P. N.: Formation of summer phytoplankton bloom in the northwestern Bay of Bengal in a coupled physical-ecosystem model, J. Geophys. Res.-Oceans, 121, 8535–8550, 2016.
Tomczak, M. and Godfrey, J. S.: Regional oceanography: An introduction, Daya Publishing House, ISBN-10: 8170353068, 2003.
Toggweiler, J. R.: Variation of atmospheric CO2 by ventilation of the ocean's deepest water, Paleoceanography, 14, 571–588, 1999.
Umling, N. E., Thunell, R. C., and Bizimis, M.: Deepwater expansion and enhanced remineralization in the eastern equatorial Pacific during the Last Glacial Maximum, Paleoceanography and Paleoclimatology, 33, 563–578, 2018.
Valley, S., Lynch-Stieglitz, J., and Marchitto, T. M.: Timing of deglacial AMOC variability from a high-resolution seawater Cadmium reconstruction: Timing deglacial upper amoc variability, Paleoceanography, 32, 1195–1203, 2017.
Van der Zwaan, G. J., Duijnstee, I. A. P., Den Dulk, M., Ernst, S. R., Jannink, N. T., and Kouwenhoven, T. J.: Benthic foraminifers: Proxies or problems? A review of paleocological concepts, Earth-Sci. Rev., 46, 213–236, 1999.
Vinayachandran, P. N., Murty, V. S. N., and Ramesh Bahu, V.: Observations of barrier layer formation in the Bay of Bengal during summer monsoon, J. Geophys. Res., 107, 8018, https://doi.org/10.1029/2001JC000831, 2002.
Xie, R. C., Marcantonio, F., and Schmidt, M. W.: Deglacial variability of Antarctic Intermediate Water penetration into the north Atlantic from authigenic Neodymium isotope ratios, Paleoceanography, 27, PA3221, https://doi.org/10.1029/2012PA002337, 2012.
You, Y.: Implications of the deep circulation and ventilation of the Indian Ocean on the renewal mechanism of North Atlantic Deep Water, J. Geophys. Res.-Oceans, 105, 23895–23926, 2000.
Yu, J., Menviel, L., Jin, Z. D., Thornalley, D. J. R., Foster, G. L., Rohling, E. J., McCave, I. N., McManus, J. F., Dai, Y., Ren, H., He, F., Zhang, F., Chen, P. J., and Roberts, A. P.: More efficient North Atlantic carbon pimp during the Last Glacial Maximum, Nat. Commun., 10, 2170, https://doi.org/10.1038/s41467-019-10028-z, 2019.
Yu, Z., Colin, C., Ma, R., Meynadier, L., Wan, S., Wu, Q., Kallel, N., Sepulcre, S., Dapoigny, A., and Bassinot, F.: Antarctic Intermediate Water penetration into the northern Indian Ocean during the last deglaciation, Earth Planet. Sc. Lett., 500, 67–75, 2018.
Zhou, X., Duchamp-Alphonse, S., Kageyama, M., Bassinot, F., Beaufort, L., and Colin, C.: Dynamics of primary productivity in the northeastern Bay of Bengal over the last 26 000 years, Clim. Past, 16, 1969–1986, https://doi.org/10.5194/cp-16-1969-2020, 2020.
Ziegler, M., Diz, P., Hall, I. R., and Zahn, R.: Millennial-scale changes in atmospheric CO2 levels linked to the Southern Ocean carbon isotope gradient and dust flux, Nat. Geosci., 6, 457–461, 2013.