the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 04 Nov 2025
| 04 Nov 2025
The most complete Holocene peat record from Central Europe: multi-proxy reconstruction of postglacial wetness changes and climate events from Linje peatland, Poland
Eliise Poolma
Katarzyna Marcisz
Leeli Amon
Patryk Fiutek
Piotr Kołaczek
Karolina Leszczyńska
Dmitri Mauquoy
Michał Słowiński
Siim Veski
Friederike Wagner-Cremer
Mariusz Lamentowicz
This study investigates the vegetation and hydrological dynamics of Linje peatland in northern Poland during the past ∼11 500 years by integrating testate amoebae and plant macrofossil analyses. The Linje profile is currently the only complete Holocene peat record in Central Europe and offers valuable insights into long-term climate variability and its ecological consequences for peatland ecosystems. The results reveal significant changes in peatland wetness and vegetation driven by autogenic processes, climatic fluctuations and anthropogenic influences. Major bryophyte species turnovers occurred ∼11 200, ∼10 350, ∼8200, 7500, ∼5500, ∼600, and ∼450 cal BP often coinciding with declines in Archerella flavum and Hyalosphenia papilio abundances. Both proxies indicate a marked period of drier conditions between ∼ 7600 and ∼ 6800 cal BP corresponding with the Holocene Thermal Maximum. Additionally, testate amoebae data suggest further disturbances at ∼ 3050, ∼ 2000, and ∼ 200 cal BP, and the latter, linked to a permanent shift in species composition, implies lasting changes to peatland conditions caused by intensified human activity. Around the end of the Early Holocene, most species turnovers and disturbances began to align with Holocene Rapid Climate Change (RCC) events.
- Article
(6426 KB) - Full-text XML
- BibTeX
- EndNote




Peatlands serve as valuable archives of past environmental conditions, capturing a record of climatic, hydrological, and ecological changes over millennia (Chambers and Charman, 2004; Karpińska-Kołaczek et al., 2024; Margielewski et al., 2024; Piilo et al., 2025; Sim et al., 2023; Swindles et al., 2019, 2025). Research on peat deposits for paleoclimate reconstruction dates back to the late 19th century, and since then methodological advances have steadily improved the accuracy, resolution, and reliability of reconstructions, enabling robust comparisons of local and global events (Chambers et al., 2012, cf. Birks and Seppä, 2010). Recent innovations include trait-based paleoecology (Marcisz et al., 2020a), biomarker analyses (Pancost, 2024), and novel techniques for quantifying wildfire intensity in peatlands (Theurer et al., 2024).
Among the various vascular plants and bryophytes that contribute to peat formation, Sphagnum mosses stand out as particularly valuable in situ biological proxies in paleoecological research (Clymo, 1984; Lamentowicz et al., 2019a; Ronkainen et al., 2014; Rydin et al., 2006). This is due to their distinct ecological preferences, physiological limitations, slow vertical growth, and resistance to decay. Different Sphagnum species are confined to specific microhabitats that vary in pH, nutrient availability, and water table depth (Rydin et al., 2006). Species turnover is strongly linked to changes in peatland hydrology, as Sphagnum species differ in their sensitivity to moisture availability in the capitulum and cannot actively regulate water uptake or prevent water loss (Gong et al., 2020). Together, these traits make Sphagnum mosses reliable indicators of past environmental conditions, particularly fluctuations in water table levels. In the Northern Hemisphere, Sphagnum mosses are major contributors to peat accumulation, especially in ombrotrophic bogs and poor fens where they typically dominate (Granlund et al., 2022; Halsey et al., 2000; Kokkonen et al., 2019; Piilo et al., 2023). In contrast, peat formation in rich fens is primarily driven by brown mosses (mainly the Amblystegiaceae family), and by the roots and rhizomes of vascular plants (Jaszczuk et al., 2024). The unique morphological, physiological, and biochemical traits of Sphagnum mosses make them effective ecosystem engineers, capable of building peat from their dead tissue while creating acidic, nutrient-poor, and waterlogged conditions. In doing so, they suppress the growth of vascular plants, further reinforcing conditions that favor their own dominance and continued growth (van Breemen, 1995; Turetsky et al., 2025). Sphagnum communities growing on sufficiently deep peat deposits are highly resilient, retaining their capacity to store carbon and recover growth after drought conditions (Lamentowicz et al., 2019a; Marcisz et al., 2020b; Moore et al., 2021).
Changes in peatland surface wetness serve as one of the indicators of past climatic shifts (Charman et al., 2009). Two of the most used proxies for detecting wet and dry shifts in peatlands are the remains of plants and testate amoebae (Blackford, 2000; Charman et al., 2006; Davies et al., 2021; Kuuri-Riutta et al., 2022; Marcisz et al., 2014a; Rundgren et al., 2023; Sim et al., 2021a; Swindles et al., 2020; Väliranta et al., 2012). While testate amoebae and plants are ecologically interconnected, testate amoebae communities can adjust species composition over shorter timescales due to their short life cycles and high turnover rates, whereas mosses and perennial vascular plants tend to respond more gradually to hydrological changes (Marcisz et al., 2014a, b; Väliranta et al., 2012). Consequently, a multi-proxy analysis of vegetation and testate amoebae assemblage dynamics provides a more comprehensive understanding of past surface wetness changes (Lamentowicz et al., 2025; Väliranta et al., 2012). Both proxies also offer insights into the trophic levels of peatlands, yielding valuable information about site development (Laine et al., 2018; Lamentowicz et al., 2010; Mauquoy and van Geel, 2007; Opravilová and Hájek, 2006).
Holocene climate variability offers an opportunity to study natural climate change before industrialization. Peat-based climatic reconstructions have mainly focused on the Northern Hemisphere spanning the Middle to Late Holocene (Charman et al., 2006; Holmquist et al., 2016; Mauquoy et al., 2008; Sillasoo et al., 2007; Swindles et al., 2010). Peatland records have also been studied for specific climate events, such as the 8.2 ka (Daley et al., 2016; Hughes et al., 2006), 4.2 ka (Jordan et al., 2017) and 2.8 ka events (Słowiński et al., 2016). In rare cases, complete Holocene records from peatlands have been identified such as Walton Moss, England (Hughes et al., 2000). Full Holocene peat records from Europe are rare because of the temporal variation in peatland succession (Ruppel et al., 2013; Stivrins, 2025) as well as the extensive history of drainage-driven land use for economic purposes (Finlayson and Spiers, 1999; United Nations Environment Programme, 2022; Verhoeven, 2014). While there are examples of full postglacial records from Polish lakes (Bonk et al., 2021; Gałka et al., 2014; Ralska-Jasiewiczowa, 1998), the longest continuous peat records reported until now came from Stążki and Głęboczek peatlands, which span 7500 and 6500 years, respectively (Gałka et al., 2013b; Lamentowicz et al., 2019b). The present study extends this record further, providing the longest continuous peat sequence documented from Poland to date. Linje in northern Poland is notable as the only known peatland in Poland with a nearly 12 m thick uninterrupted peat layer that covers the past ∼ 11 500 years of its development history. Various research projects have been conducted on the site exploring its history, current biotic and abiotic environment. Open-top chambers on the site have been used to model climate change effects on local vegetation, plant-fungal interactions, microbial activity and peatland respiration processes (Buttler et al., 2023; Jassey et al., 2018; Reczuga et al., 2018; Samson et al., 2018). Additional research papers have explored the hydrological functioning and microclimatic properties of Linje (Słowińska et al., 2010, 2022) and testate amoebae seasonal changes in response to hydrology (Marcisz et al., 2014b). Research by Marcisz et al. (2015) has focused on the high-resolution hydrological dynamics and fire history at the site over the last 2000 years, whereas Szambelan et al. (2025) focused on the human impacts on the surrounding landscape between approximately 3220 and 1550 cal BP. Earlier paleoecological investigations covering the full developmental history of Linje peatland include the works of Noryśkiewicz (2005), Kloss (2005) and Kloss and Żurek (2005). However, these studies were largely preliminary, offered limited or no possibilities for quantitative analysis, and lacked cross-validation from multiple proxies.
In this study, we investigate a nearly 12 m long undisturbed peat core from Linje peatland. To our knowledge, it is the only continuous peat core in Central Europe that reflects the environmental dynamics of the entire Holocene (11 500 years). Our analysis centres on plant macrofossil and testate amoebae analysis, particularly emphasising changes in the species communities of bryophytes and testate amoebae. We aim to reconstruct local vegetation and wetness changes while exploring their connections with Holocene climatic fluctuations to better understand past ecosystem responses to climate change.
The studied core was extracted in 2019 CE from Linje ( N, E), a 5.95 hectare peatland located near Bydgoszcz in northern Poland (Fig. 1). Situated within the nemoral-continental vegetation zone (Preislerová et al., 2024), the Linje peatland forms part of the 12.7 ha Linje Nature Reserve, established in 1956 (Kucharski and Kloss, 2005). The surrounding landscape is a legacy of the last glacial and the peatland formed in a ∼12 m deep depression (Żurek, 2005).
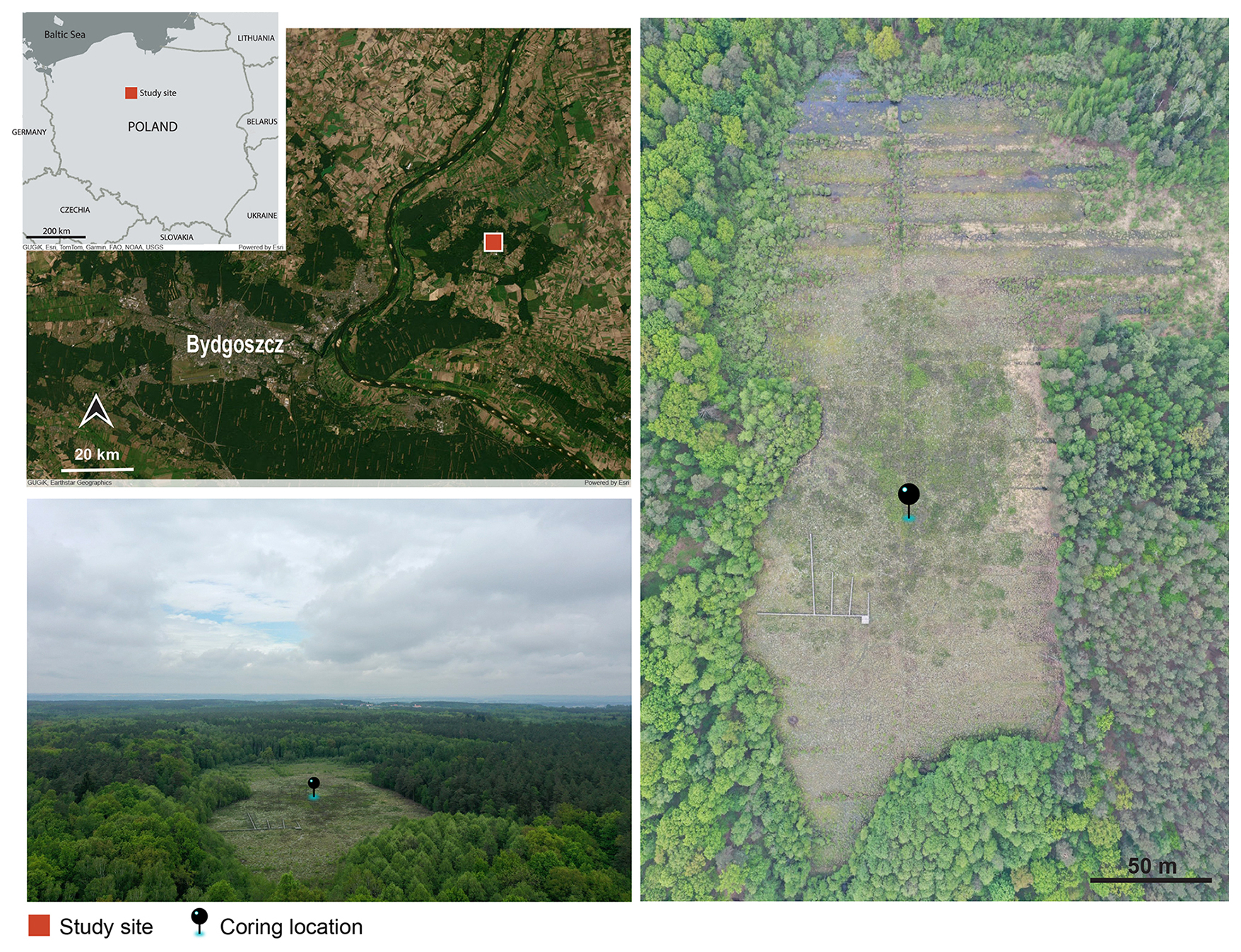
Figure 1Location of the Linje peatland study site. Basemaps (top left) are from ArcGIS Online (modified), photographs (right and bottom left) by Michał Słowiński.
The peatland is surrounded by a mosaic of mixed forests (Betula pendula, Quercus robur, Pinus sylvestris, Picea abies), arable land and villages (Kucharski and Kloss, 2005). Recognized as a microrefugium for protected dwarf birch (Betula nana), the site is protected under the Natura 2000 network (Marcisz et al., 2015; Noryśkiewicz, 2005).
The site features a gradient of vegetation types, ranging from poor fen at the periphery to ombrotrophic conditions in the centre reflecting its hydrological functioning (Noryśkiewicz, 2005; Słowińska et al., 2010). Since Linje is a small, forest-surrounded peatland in a depression, it has a cooler microclimate than a nearby open-air reference site (Słowińska et al., 2022). The authors point out that 19th-century drainage efforts have made the peatland surface highly responsive to meteorological changes, contributing to hydrological instability. This instability has also been observed in a testate amoeba-based water table reconstruction (Marcisz et al., 2015).
3.1 Coring and radiocarbon dating
In 2019 two parallel peat profiles with a distance of 50 cm were cored from the centre of Linje peatland with a Russian type corer (chamber dimension: 10 cm×100 cm), from a vegetation area characteristic of the ombrotrophic section of the site. The longer profile (Linje_2019 core; Szambelan et al., 2025), with a total length of 1230 cm, was chosen for analyses in this study and it was cut into 2.5 cm thick sections.
Radiocarbon dating was performed on selected plant macrofossils (Table 1) at the Poznań Radiocarbon Laboratory (Poland) using their standard pre-treatment protocols. The chronology of the peat profile is based on a Bayesian age-depth model generated from 12 out of 14 14C AMS dates (Table 1). The model was constructed using OxCal. 4.4.4 software (Bronk Ramsey, 1995, 2006), by applying the P_Sequence function with the following parameters: k0=0.75, and interpolation=0.5 cm (Bronk Ramsey, 2008; Bronk Ramsey and Lee, 2013). The IntCal20 (Reimer et al., 2020) 14C atmospheric curve was used as the calibration set. Dates Poz-128079 and Poz-128073 were distinctly younger than the neighbouring dates. Preliminary model runs, both with the full set of dates included, and with only one problematic date excluded, showed that these two ages had very poor agreement with the model. Therefore, both were excluded from the final age-depth model. The sections of the profile in which peat properties point to potential changes in peat accumulation rate (ARpeat) were introduced to the model as boundaries (Boundary command). These were placed at depths: (i) 1230 cm – the base of the model, (ii) 1130 cm – transition from higher water tables towards Sphagnum peat, retrieved from low-resolution palynological data (data not published) and plant macrofossils (iii) 0 cm – top of the profile.
Table 1AMS radiocarbon dates from the Linje_2019 profile. Poz = Poznań Radiocarbon Laboratory.
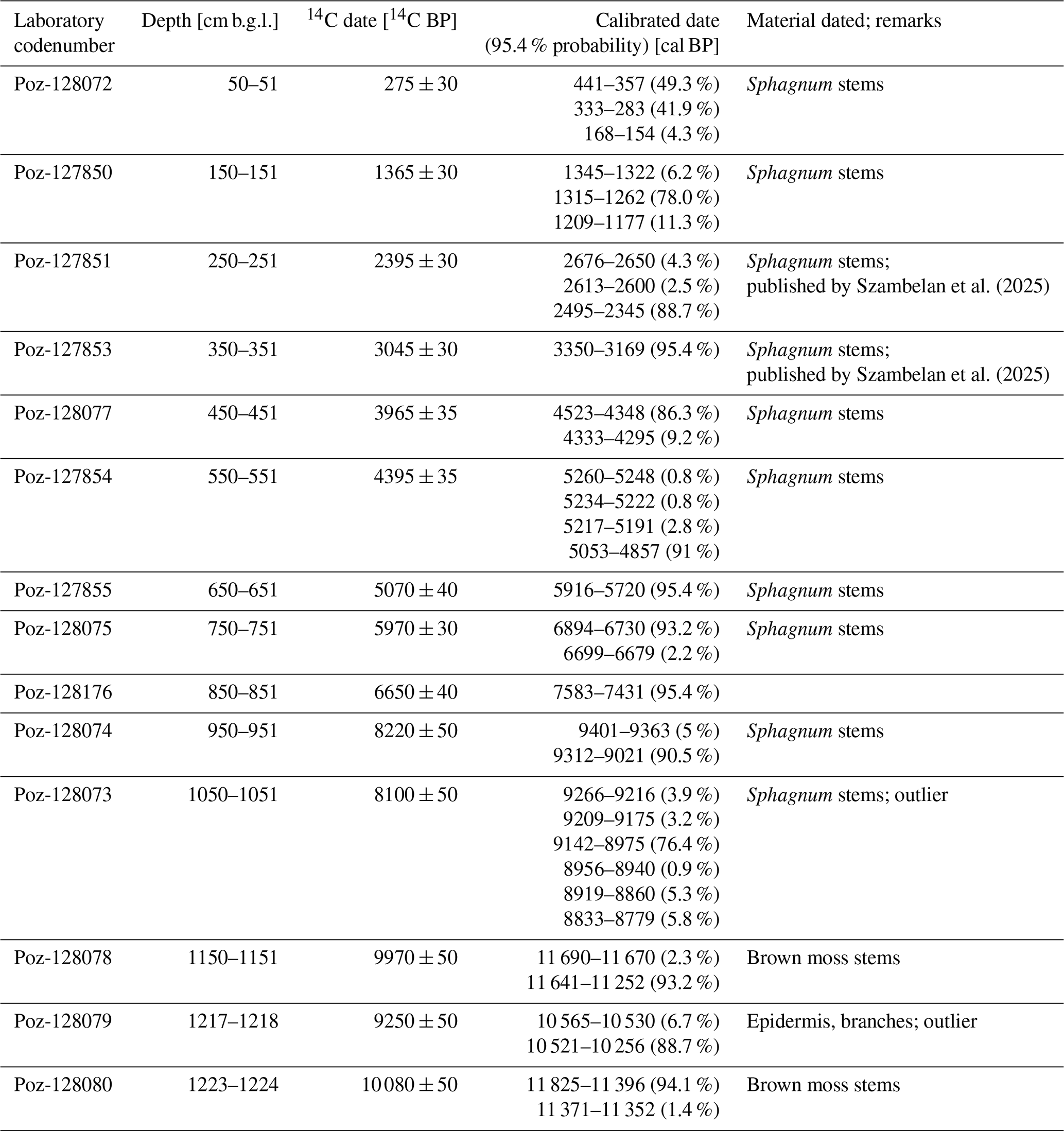
In the following sections of this article, μ (mean) values retrieved from the age-depth model were rounded to the nearest 50 years. The age was expressed as cal BP (before present).
3.2 Plant macrofossil analysis
Plant macrofossils were analysed in subsamples (volume: 2–27 cm3) with a 5–10 cm resolution from depths 2.5–1187.5 cm. The analysed volume depended on the available sediment. The section 1187.5–1230 cm was not analysed to keep these data consistent with testate amoebae as below 1187.5 cm testate amoebae were not present in significant numbers. Samples were sieved through a 125 µm mesh and disaggregated using tap water without chemical pre-treatment. The residue on the sieve was observed under a stereomicroscope (Olympus SZX9) and a light microscope (Nikon Eclipse 90i). Quadrat and Leaf Count protocols (Barber et al., 1994; Mauquoy et al., 2010) were used to estimate the peat components and Sphagnum species compositions in percentages. When total Sphagnum was ≤5 % of the sample then the Sphagnum species were not identified. Seeds, bud scales, catkin scales etc. were counted as absolute numbers and calculated as concentrations for 5 cm3. During Sphagnum identification the branch leaves having most similarity to S. angustifolium, S. balticum and S. fallax were marked down as “Sphagnum recurvum complex” (Duffy et al., 2020). Stem leaves of all these species were found and counted separately. Other species that are difficult to identify in the fossil form include S. divinum/medium and S. rubellum/fuscum, thus they are combined in the macrofossil diagram. The presence of S. fuscum in Sphagnum section Acutifolia-dominated samples was confirmed with stem leaves and dark stem colour. Sphagnum identification followed (Laine et al., 2018) and a reference collection. Brown mosses and vascular plant remains were identified using several keys (Cappers et al., 2012; Grosse-Brauckmann, 1972; Katz et al., 1977; Mauquoy and van Geel, 2007; Tomlinson, 1985) and a reference collection. Vascular plant and bryophyte nomenclature follows GBIF Backbone Taxonomy (GBIF Secretariat, 2023).
In addition to botanical remains, the percentages of unidentified organic matter (UOM) and macrocharcoal (≥1 mm) were estimated in the samples as a part of the Quadrat method.
3.3 Testate amoebae analysis
Testate amoebae were analysed in ca. 5 cm3 subsamples with a 5 cm resolution from depths 0–1187.5 cm. Samples were washed under a 300 µm mesh following the method described by Booth et al. (2010). Testate amoebae were analysed under a light microscope with ×200 and ×400 magnifications until a sum of 150 tests per sample was reached (Payne and Mitchell, 2009). In a few layers where the concentration of individuals was lower, testate amoebae were counted until a sum of 100 or 50 was reached; these were still suitable for performing further statistical analyses (Payne and Mitchell, 2009). To achieve the highest taxonomic resolution, species identifications were supported by several taxonomic keys and monographs (Clarke, 2003; Mazei and Tsyganov, 2006; Meisterfeld, 2000, 2001; Ogden and Hedley, 1980), and online resources (Siemensma, 2024). The results were used to reconstruct palaeohydrological conditions at the site.
3.4 Statistical analyses and data presentation
The stratigraphical diagrams were created in Tilia v. 3.0.1 (Grimm, 2011) or C2 v. 1.8.0. (Juggins, 2007) and edited in Adobe Illustrator (Adobe Inc., 2019). Subfossil bryophytes were used as a main proxy of this article, thus the changes on the peatland are based on the changes in bryophyte assemblages. Sphagnum leaf count data and the abundance of other mosses were extracted from the general plant macrofossil data, and the percentages were re-calculated considering the taxonomic proportions of bryophytes only. These bryophyte data were analysed in RStudio (R Core Team, 2024) to create a stratigraphic zonation. The vegdist function from the vegan package (Oksanen et al., 2001) was used to compute a Bray-Curtis dissimilarity matrix from the bryophyte data, then hierarchical clustering was performed on the dissimilarity matrix using the constrained incremental sum of squares (CONISS) from the rioja package (Grimm, 1987; Juggins, 2009). The broken-stick method was used to determine the number of meaningful clusters (Bennett, 1996). Initially 11 clusters were identified. However, cluster 8 fell below the reference line (Fig. A1), Thus, cluster 8 was merged with cluster 7, resulting in a final solution of 10 clusters. In addition, bryophytes were grouped according to their hydrological microhabitat preferences (Table A1). The percentages of wet microhabitat species were transformed to negative values following the example of Piilo et al. (2023), so that the summed taxon percentages yield a bryophyte-based moisture curve. The results of a testate amoebae analysis were used for the quantitative depth-to-water table (DWT) reconstruction, which was performed in RStudio (R Core Team, 2024) using the Pan-European training set (Amesbury et al., 2016). Testate amoebae and bryophyte species composition (total 48) were also visualised as a Non-Metric Multidimensional Scaling (NMDS) ordination plot across different samples (total 178) using RStudio and the metaMDS function in the vegan package. To calculate dissimilarity between sites, the Bray-Curtis distance metric was used. For illustrating disturbances to the peatland, the abundance of Sphagnum and the sum of abundances of Archerella flavum and Hyalosphenia papilio species were plotted. These testate amoebae species were selected as they were dominant in the Linje core and considered as indicators of wet and stable conditions on peatlands (Łuców et al., 2022; Marcisz et al., 2014a). Thus, the drop in their abundance would suggest a disturbance.
4.1 Chronology
The age-depth model (Fig. 2) recorded a model agreement (Amodel) equal to 66.7 %, which is above the recommended minimum of 60 % (Bronk Ramsey, 2008). The profile spans the time period between cal BP (2019 CE) and (1σ error) cal BP. The mean 1σ error was ca. 65 cal years. For the topmost 100 cm we lack the chronological control to detect short-term differences in accumulation rates. This section should thus be interpreted with caution due to this limitation as it is based on one date and it does not reflect an exponential age-depth relationship as it should with well-developed acrotelm and catotelm layers (Marcisz et al., 2015).
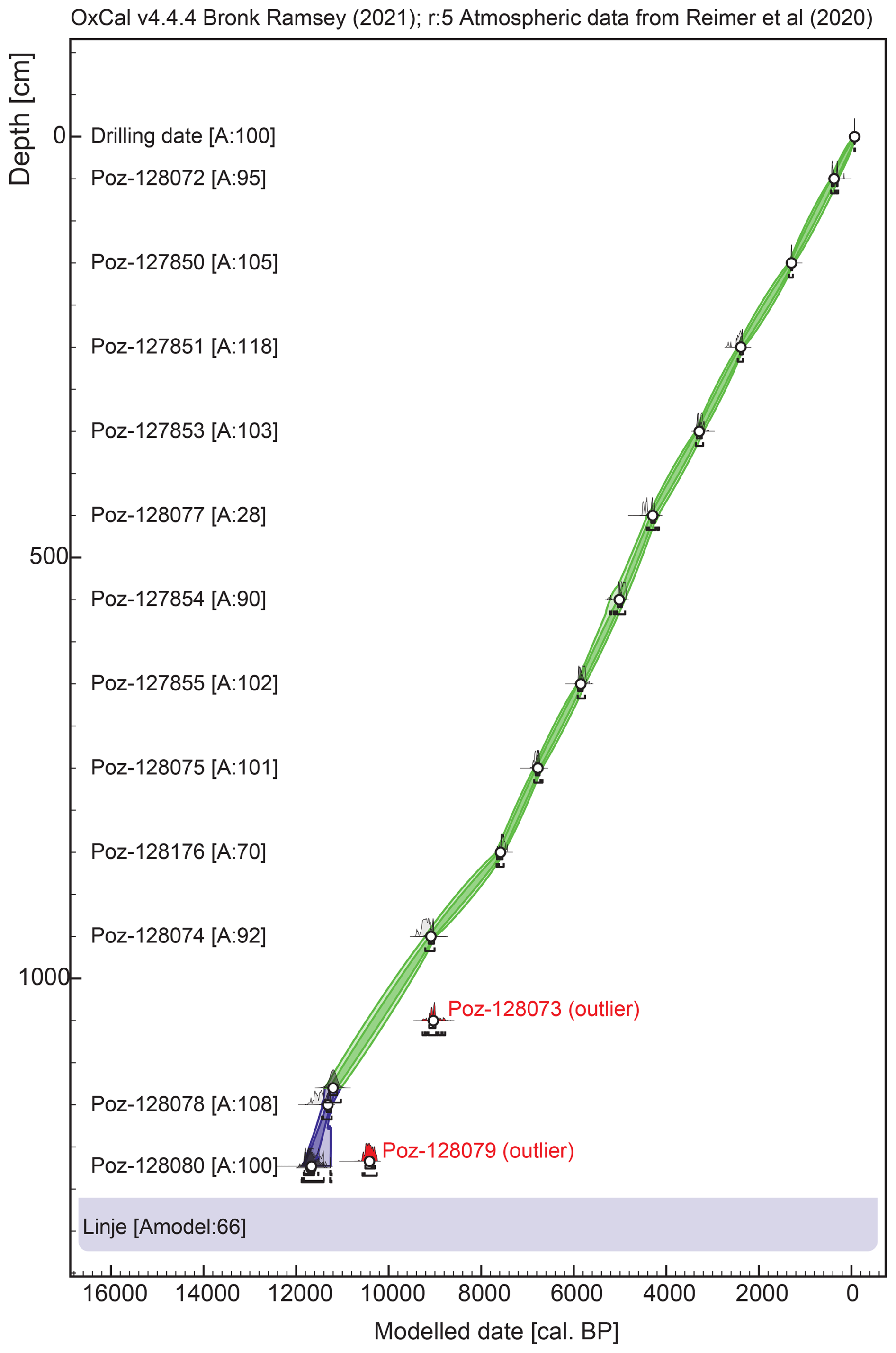
Figure 2Bayesian age-depth model of the Linje core based on 20 radiocarbon dates (Table 1). The model was constructed using OxCal. 4.4.4 software (Bronk Ramsey, 1995, 2006).
4.2 Environmental changes
Ten zones were identified based on statistically significant changes in the bryological assemblages (Figs. A1 and 3). These zones, along with general plant macrofossil dynamics and testate amoebae assemblages (Figs. 4 and 5), are discussed in detail below. Depth to water table (DWT) reconstructions are derived from testate amoebae analyses.
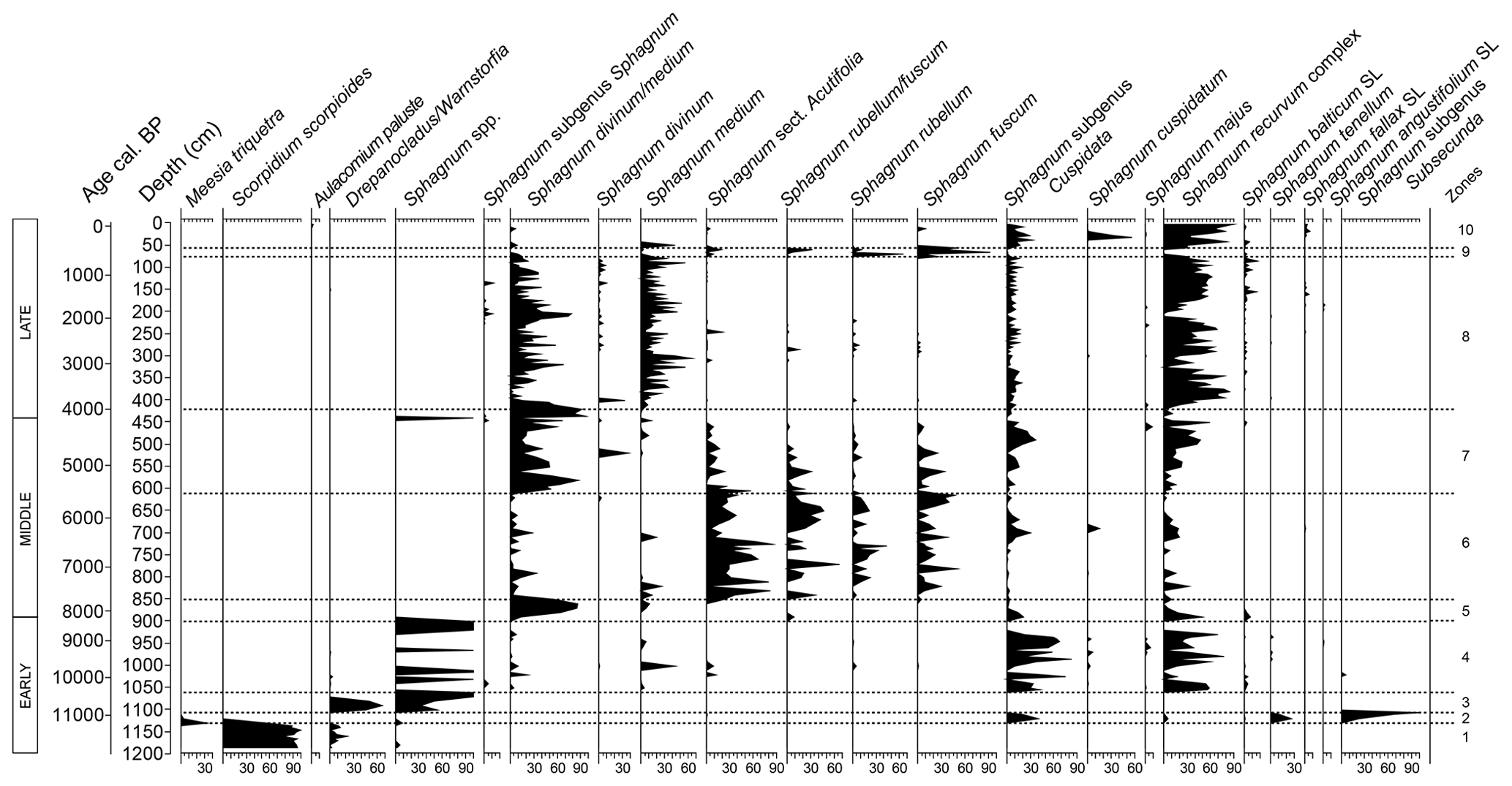
Figure 3Bryophyte stratigraphy (%) from the Linje peatland core throughout Holocene. Percentages (black) were recalculated from the general plant macrofossil dataset based on bryophyte taxonomic proportions to establish core zonation in R Studio. SL = stem leaves.
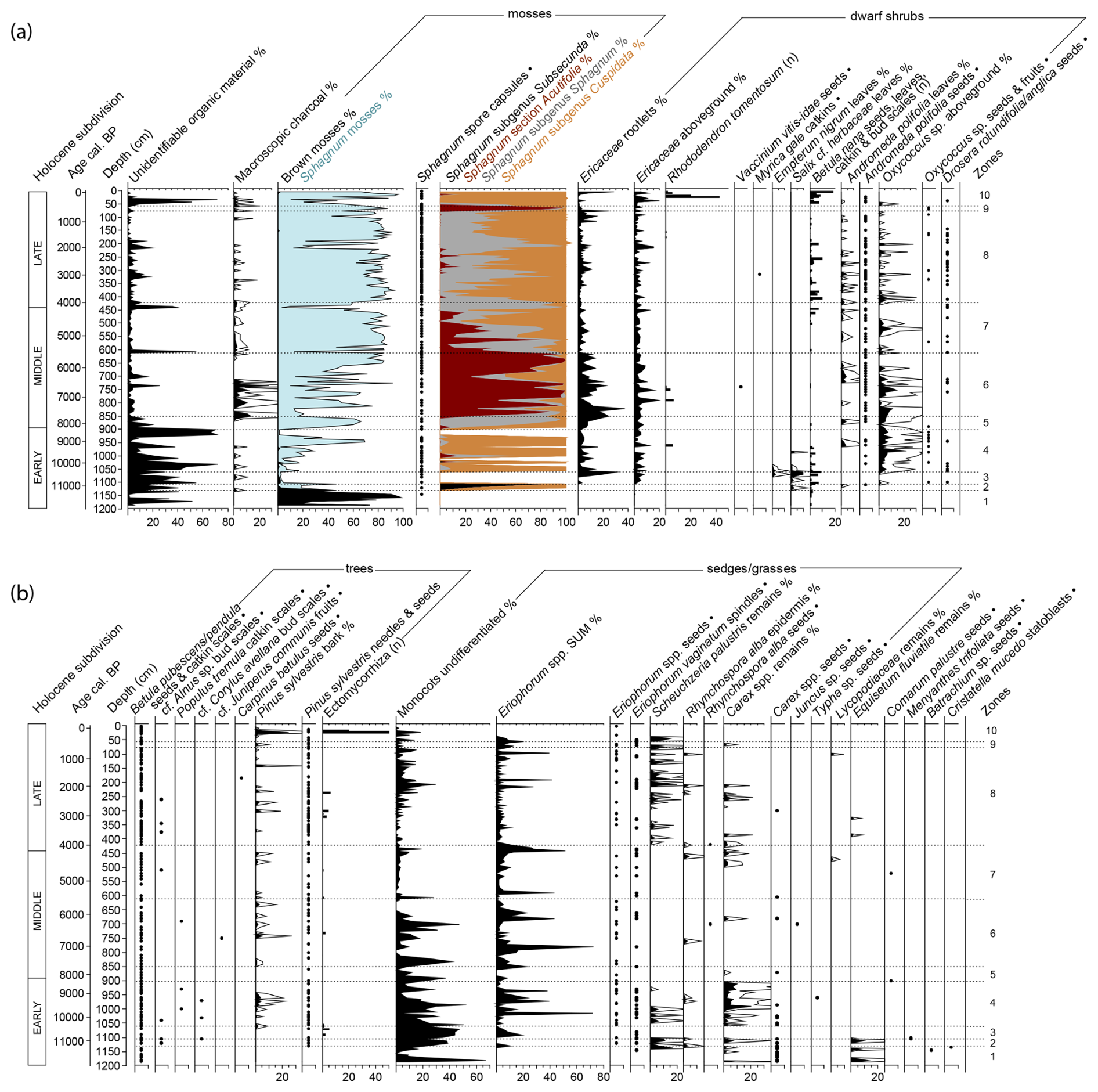
Figure 4(a, b) Plant macrofossil stratigraphy from the Linje peatland core throughout Holocene. Histograms (black or coloured) represent taxon percentages with ×5 exaggeration (when needed). Abundance of countable macrofossils (n) is shown in bars, and presence is indicated by dots (•). Zonation (dashed line) follows bryophyte assemblages (see Fig. 3). (a) includes sum of Sphagnum moss proportions, grouped by subtaxa and differentiated by colour.
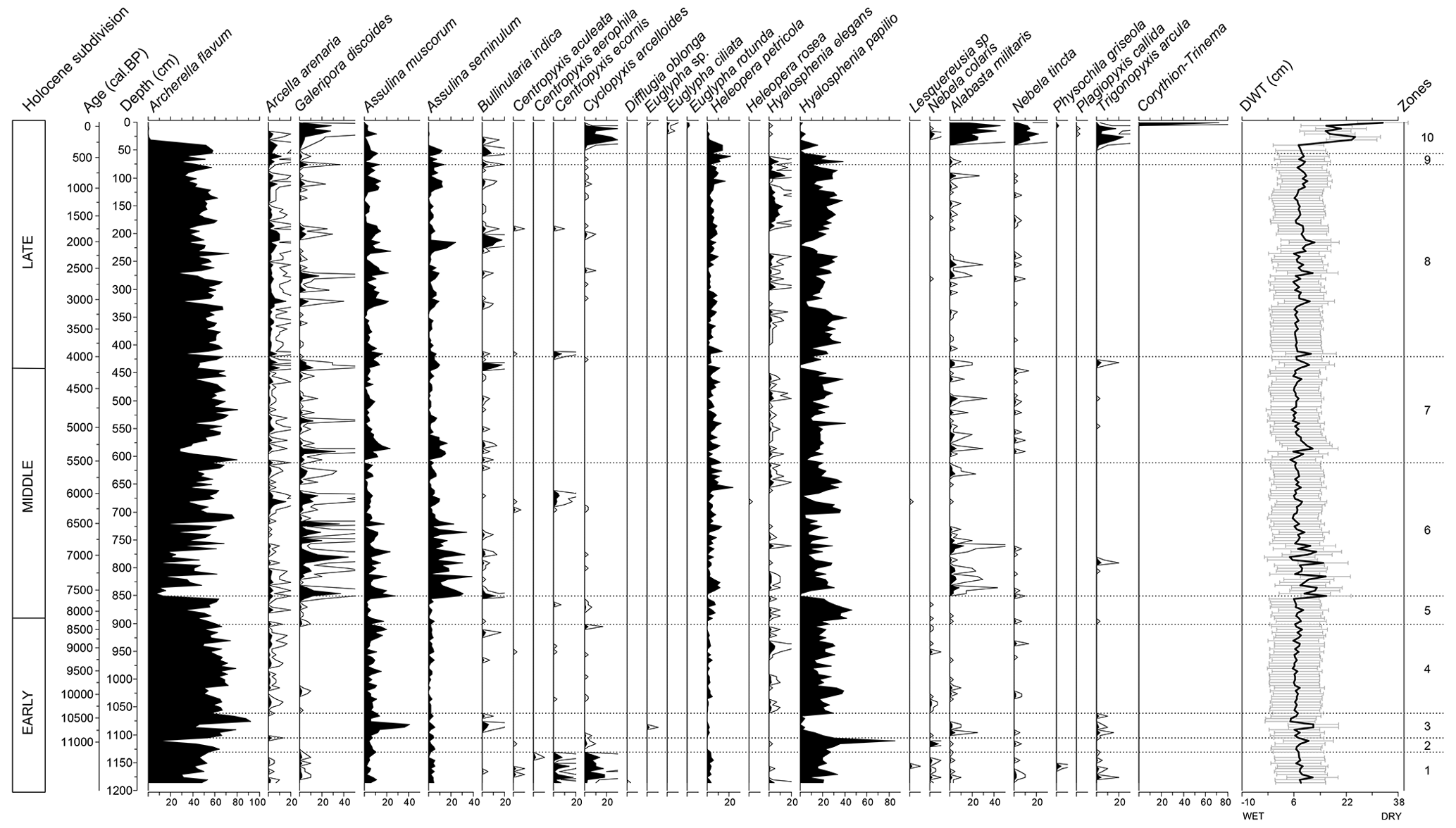
Figure 5Testate amoebae stratigraphy (%) from the Linje peatland core throughout the Holocene. Where necessary, curves are exaggerated 5× for better visibility, The depth-to-water table (DWT, cm) curve illustrates the reconstruction derived from the testate amoebae assemblages. Zonation (dashed line) follows the bryophyte assemblages (see Fig. 3).
The Linje peatland macrofossil record (Fig. 4) is abundant in bryophytes, which constitute the largest proportion of peat-forming components. At the beginning of the study's timeline, the moss assemblage (Fig. 3) reflected high water levels, nutrient availability, and elevated pH. After ∼11 200 cal BP, brown mosses were gradually replaced by Sphagnum species, coinciding with the first appearances of typical oligotrophic mire plants such as Andromeda polifolia, Oxycoccus sp., and Drosera spp. Around 10 350 cal BP, a decline in pH and nutrient levels is evident as species from the Sphagnum recurvum complex established dominance, which persisted until ∼8200 cal BP, when Sphagnum divinum/medium became the prevailing taxa. Approximately 400 years later (∼7500 cal BP), Sphagnum sect. Acutifolia began to dominate until ∼5500 cal BP, when the interplay between S. divinum/medium and Sphagnum recurvum complex species began. Sphagnum sect. Acutifolia experienced only a brief resurgence ∼600–450 cal BP before being replaced by Sphagnum subgen. Cuspidata species, which have remained dominant to the present.
From the plant macrofossil record it is evident that birch (Betula pubescens/pendula) was already growing near Linje peatland ∼11 500 cal BP. Pinus sylvestris macrofossils were first found in the record at ∼11 200 cal BP. Wetter periods are marked by increased abundances of Equisetum fluviatile, Carex spp. and Scheuchzeria palustris macrofossils, whereas drier intervals are sometimes characterized by a higher proportion of ericaceous remains.
Testate amoebae-based water table reconstruction indicates that over the past 11 300 years, the average depth to the water table was 7.6 cm, increasing to a maximum of 15 cm during the Middle Holocene (Fig. 5). Depths exceeding 16 cm occurred only in the past ∼200 years. Since the beginning of the record, the dominant testate amoebae species at the site have been A. flavum and H. papilio. After ∼7500 cal BP declines in the abundance of A. flavum and H. papilio closely correspond with decreases in Sphagnum abundance. The two proxies exhibit strong general agreement in hydrological interpretation, with the exception of the last 600 years.
In this study, “disturbance” refers to phases of hydrological instability. At Linje, these are marked by elevated proportions of unidentified organic material (UOM), reduced test counts, and declines in dominant testate amoebae taxa (A. flavum, H. papilio), as well as reductions in Sphagnum abundance once the peatland became Sphagnum-dominated after ∼9050 cal BP. Sudden peaks in UOM are interpreted as reflecting periods of increased peat decomposition, which are associated with peat surface drying in otherwise stable peatland systems (Loisel et al., 2017; Sim et al., 2021b). Disturbance episodes are also associated with higher abundances of testate amoebae species such as Galeripora discoides, Assulina muscorum, and Assulina seminulum, which are linked to either drier conditions (Assulina spp.) or fluctuating water tables (G. discoides) (Lamentowicz et al., 2008b). Despite multiple disturbance episodes, major testate amoebae species turnover is absent until a shift ∼200 years ago. Importantly, these disturbances were relatively short-lived, and peat accumulation resumed quickly, resulting in a stable and continuous age-depth model.
Zone 1: ∼ 11 500–11 200 cal BP (1187.5–1130 cm) – waterlogged, rich fen
Bryophytes are dominated by Scorpidium scorpioides and some Drepanocladus/Warnstorfia-type mosses are also present. In samples dated to ∼11 200 and ∼11 100 cal BP macrofossils of Meesia triquetra were also found. S. scorpioides and M. triquetra, considered characteristic boreal fen species (Hájková et al., 2015), are found in many Late glacial and Early Holocene sediments in northern Poland, but their present status in Central Europe is rare and endangered (Drzymulska, 2024; Hájková et al., 2015; Słowiński et al., 2015, 2017). Betula sect. Albae seeds are present from the beginning of the record, as well as Betula nana leaves, bud scales and seeds indicating that dwarf birch was growing on the site and tree-birches were already present in the region. Around 11 300 cal BP, the presence of Eriophorum vaginatum is indicated by a spindle, although epidermis of this species was not found. Carex spp. remains and nutlets are found in this zone, as well as E. fluviatile remains. Around the end of zone 1 (∼11 300 cal BP) a seed of water-crowfoot (Batrachium sp.) was found, followed by a statoblast of Cristatella mucedo (freshwater sponge) around 11 250 cal BP which indicate inundated conditions or a shallow water habitat (Lumbreras et al., 2011; Økland and Økland, 2000). S. palustris remains also appear at the end of the zone, strengthening the interpretation of high water tables or the presence of a standing water pool (Tallis and Birks, 1965).
The testate amoebae assemblage in this zone is dominated by A. flavum and H. papilio, which are associated with wetter microhabitats (Lamentowicz and Mitchell, 2005; Marcisz et al., 2014a). Centropyxis ecornis is present throughout this zone and disappears at the end of this phase. According to a Polish training set, this species indicates wet and nutrient-rich conditions (Lamentowicz et al., 2008a). Another distinctive feature of this zone is the presence of Cyclopyxis arcelloides, which is usually found in wet habitats in Poland (Lamentowicz and Mitchell, 2005). Both C. ecornis and C. arcelloides, as well as Trigonopyxis arcula that were present in this phase, incorporate mineral material in their tests which can indicate a source of mineral inputs into the peatland (Marcisz et al., 2020a). The abundance of species indicating drier and more acidic conditions such as A. seminulum and A. muscorum (Lamentowicz et al., 2008a) remains under 20 %. Reconstructed water table depths average at 8.3 cm.
Zone 2: ∼ 11 200–10 900 cal BP (1130–1105 cm) – wet, rich fen
Although not dominant, the Sphagnum assemblage indicates rich fen conditions with S. cuspidatum, S. tenellum and Sphagnum from subgenus Subsecunda (mainly S. contorum) (Gałka and Lamentowicz, 2014; Laine et al., 2018; Mauquoy and van Geel, 2007). In this zone, the first macrofossils (needles, bud scales and seeds) of Pinus sylvestris and a bud scale of cf. Alnus sp. were found, indicating their presence in the wider landscape. E. vaginatum epidermis and seeds were also found for the first time at the beginning of this zone. Vascular plants indicate persistently wet conditions with abundant Carex spp., E. fluviatile and S. palustris remains. Moreover, seeds of Menyanthes trifoliata were identified in samples dated to ∼10 900 and ∼10 850 cal BP. In a sample dated to ∼10 900 cal BP a bud scale of cf. Corylus avellana was found. Sphagnum mosses increase in abundance between ∼11 100 and ∼11 000 cal BP with ombrotrophic to weakly minerotrophic Sphagnum tenellum (Laine et al., 2018) and Sphagnum subgen. Cuspidata.
The dominant testate amoebae species are still A. flavum and H. papilio. Both species increase and become more numerous in this zone. C. ecornis and Heleopera petricola disappear during this phase and the abundance of C. arcelloides decreases. Nebela collaris is relatively abundant at this stage. Assulina species occurrence does not change significantly compared to the previous zone. Reconstructed water table depths are similar to the previous phase, pointing to wet conditions.
Zone 3: ∼ 10 900–10 400 cal BP (1105–1060 cm) – a shift towards drier and nutrient poor conditions
Although not abundant, typical oligotrophic species such as Oxycoccus palustris and Drosera rotundifolia/anglica are found for the first time at this stage. Differentiating between the seeds of the two Drosera species is difficult. D. rotundifolia tends to indicate lower pH and exhibits a broad ecological tolerance, whereas D. anglica is typically associated with wetter conditions and slightly higher pH levels (Banaś et al., 2023). Nonetheless, the presence of either insectivorous species implies increasing nutrient limitation in the peatland environment (Mauquoy and van Geel, 2007). After ∼10 900 cal BP, when Sphagnum subgen. Subsecunda was relatively abundant, Sphagnum mosses decline. Since they occupied less than 5 % of the peat components, species were not identified. A very small proportion of Drepanocladus/Warnstorfia-type brown moss is present. The main peat components are UOM, ericaceous rootlets and undifferentiated monocots. The shift towards drier conditions is seen in plant macrofossils since ericaceous remains appear in this stage, including arctic/boreal shrubs Salix cf. herbacea and Empetrum nigrum, that are more abundant during the end of this phase.
The conditions on the peatland remain wet, as the testate amoebae-inferred water table averages 7.6 cm, however slightly drier conditions occur between ∼10 700 and ∼10 650 cal BP when the water table depth is around 12 cm. This is coupled with significant changes in the species composition: a reduction in the abundance of H. papilio is followed by a slight increase of A. muscorum and Bullinularia indica, the latter two species being associated with drier conditions (Lamentowicz et al., 2008a).
Zone 4: ∼ 10 400–8350 cal BP (1060–900 cm) – moderately wet phase, poor fen
The plant macrofossil record indicates wet conditions with abundant S. palustris, and Carex spp. remains (including nutlets) as well as some fragments of Rhynchospora alba stem. From ∼10 200 cal BP onwards E. nigrum does not appear in the plant macrofossil record. S. cf. herbacea remains occur occasionally during this period, lastly in a sample dated to ∼9500 cal BP. Sphagnum mosses start to be more present again after ∼10 350 cal BP. In the following samples, wet habitat species: S. majus, S. cuspidatum and especially species from the Sphagnum recurvum complex (stem leaves of S. balticum and S. angustifolium) were found. The Sphagnum stratigraphy is interrupted by a disturbance ∼8500–8350 cal BP characterized by a large proportion of UOM. At ∼9700 cal BP the presence of Populus tremula is indicated with a catkin scale.
The testate amoebae community shows this period as stable and wet. Compared to the previous phase, the abundance of wet indicator species, H. papilio and A. flavum increase and remain abundant until a slight drop around 8500 cal BP.
Zone 5: ∼ 8350–7600 cal BP (900–850 cm) – a shift to drier conditions, poor fen
After a disturbance in the previous phase, this phase is characterized by a shift in Sphagnum composition towards drier conditions. Previously dominating wet-habitat Sphagnum subgen. Cuspidata is replaced by S. divinum/medium, which might suggest development of a drier microform (Blackford, 2000; Hassel et al., 2018).
The testate amoebae community composition does not show significant changes during this time. The abundance of H. petricola and H. papilio increases slightly, while A. muscorum abundance values drop.
Zone 6: ∼ 7600–5550 cal BP (850–610 cm) – dry phase, poor fen
Around 7600 cal BP, a notable decrease in the general abundance of Sphagnum is observed, coinciding with the first appearance of Sphagnum sect. Acutifolia species, primarily S. rubellum and S. fuscum, in the samples. By approximately 7500 cal BP, Sphagnum sect. Acutifolia begins to dominate over S. divinum/medium. Both S. rubellum and S. fuscum typically inhabit hummocks, which are more drought-resistant than hollows (Mauquoy and van Geel, 2007; Nungesser, 2003).
During this period, the samples are notably rich in macroscopic charcoal (≥1 mm), which constitutes 10 %–13.5 % of the peat at the onset of this zone (∼ 7600–7500 cal BP). Similar levels of macrocharcoal abundance are detected at approximately 7200 and 6800 cal BP. By around 6650 cal BP, macro-charcoal pieces account for more than a quarter of the sample (27.6 %), which is accompanied by a decrease in Sphagnum (9.7 %) and an increase in the percentage of UOM (25.8 %). Macrocharcoal-rich samples, with minor interruptions, persist until about 6500 cal BP, correlating with an increase in aboveground ericaceous remains and the presence of drier habitat Sphagnum species. Given the limited transport distance of macrocharcoal (Clark, 1988; Peters and Higuera, 2007; Pisaric, 2002; Tinner et al., 2006), these phases likely reflect local fire activity.
The second half of this zone, beginning around 6400 cal BP reveals a transition toward moderately wet conditions, evidenced by the increased occurrence of Sphagnum subgen. Cuspidata and remains (including seeds) of R. alba and Carex spp. These changes indicate increased peatland surface wetness between ∼ 6300 and ∼ 6150 cal BP. B. nana remains were not present in the entire zone, possibly because the drier microform conditions were not optimal for preserving the remains.
Testate amoebae-based water table reconstructions indicate that the water table was significantly lower at the start of this period with an average of 10 cm between ∼ 7600 and ∼ 6800 cal BP. The abundance of dominating species A. flavum and H. papilio declines sharply around 7600 cal BP but quickly recovers to previous high levels. This drop and generally low counts of testate amoebae may be linked to a local fire event, as the sample was also rich in macro-charcoal. A similarly low count of testate amoebae is observed in a sample dated to ∼7050 cal BP, which is low in Sphagnum and particularly rich in E. vaginatum remains. In the initial phase, Assulina spp. and A. militaris are abundant, while the highly fluctuating abundance of G. discoides is characteristic of the entire zone. Lamentowicz et al. (2008a, b) note that this species can indicate fluctuating water tables, sometimes linked to disturbance. The DWT reconstruction shows a transition to wetter and more stable conditions starting from ∼ 6800 cal BP.
Zone 7: ∼ 5550–4000 cal BP (610–420 cm) – a shift to wetter conditions, poor fen
This phase is wetter than the previous one, as indicated by a more heterogeneous Sphagnum composition: S. divinum/medium and the S. recurvum complex occur in large proportions, while Sphagnum sect. Acutifolia species are still present at the beginning of the phase but gradually decline thereafter. A peak of Sphagnum subgen. Cuspidata occurred ∼ 4600 cal BP. This is followed by the occurrence of Carex spp. and R. alba in the plant macrofossil record, indicating that the second half of the phase is wetter. B. nana, which was absent from the record for approximately 1000 years, is present again from ∼5450 cal BP onwards. The UOM that mainly remained less than 5 % of the sample since ∼6250 cal BP, reaches 55 % ∼ 5500 cal BP and 36 %–40 % ∼ 4200–4150 cal BP. In these samples, Sphagnum abundance drops. The sample dated to ∼4200 cal BP also has a very high count (>100) of E. vaginatum spindles and a high percentage of E. vaginatum roots. The following younger samples also contain aboveground epidermis of E. vaginatum, which could suggest a period of hydrological instability (Barber et al., 2004; Gałka et al., 2017a; Łuców et al., 2022).
The testate amoebae species composition is similar to the previous zone. Both disturbance events are also shown in the testate amoebae assemblage – test count is low around ∼5500 and ∼4200–4100 cal BP. During the ∼5500 cal BP disturbance the abundance of H. papilio decreases abruptly, while the abundances of A. flavum decrease slightly resulting in a DWT of 5 cm. During the disturbance ∼4210–4110 cal BP, abundances of both species decrease abruptly and recover quickly acting similarly to Sphagnum mosses. DWT remains 7.8 cm during ∼ 4200 cal BP, but averages to 10 cm by ∼ 4100 cal BP.
Zone 8: ∼ 4000–600 cal BP (420–75 cm) – wet phase, poor fen
Sphagnum mosses indicate wet conditions with a general dominance of Sphagnum subgen. Cuspidata. Numerous S. balticum stem leaves were found, although S. fallax and S. angustifolium stem leaves were also present. S. divinum/medium is also present throughout this zone, with a peak abundance ∼ 3050 and ∼ 1950–1900 cal BP. Sphagnum sect. Acutifolia remains between 0 %–23 % throughout this phase, being the highest ∼ 2350 and ∼ 2700 cal BP. This zone shows a strong presence of S. palustris and the occasional presence of Carex spp., R. alba and E. fluviatile – all plants that grow in wet conditions. These vascular plants indicate especially wet conditions ∼ 3750–3650, ∼ 2500–2300 and ∼ 1950 cal BP. The Sphagnum percentage drops to 12 % ∼2000 cal BP and remains low until ∼1900 cal BP. In a sample dated to ∼1700 cal BP a Carpinus betulus seed was present.
The testate amoebae record indicates that this period was moderately wet. The abundance of Hyalosphenia elegans increases in the samples during the whole zone, which confirms wet conditions (Lamentowicz and Mitchell, 2005). Disturbance on the peatland occurs at ∼3050 cal BP indicated by decrease in abundance of A. flavum and H. papilio. Around 2000 cal BP, drier conditions are inferred from an increase in B. indica and A. seminulum abundances, accompanied by a decline in H. papilio. This shift corresponds to a reconstructed depth to water table drop to approximately 12 cm. This period also coincides with low Sphagnum percentages and an increase in decomposed organic matter, collectively indicating increased hydrological instability.
Zone 9: ∼ 600–450 cal BP (75–55 cm) – dry phase, poor fen
This ∼200 year-period is characterized by a sudden peak in Sphagnum sect. Acutifolia (mostly S. fuscum) which replaces moderately wet habitat species from the previous phase. The testate amoebae and vascular plant record do not show any changes compared to the previous phase.
Zone 10: ∼ 450 to −69 cal BP (2019 CE) (55–0 cm) – water table fluctuations
The Sphagnum composition switches from S. fuscum (sect. Acutifolia) to wet habitat Sphagnum species – S. cuspidatum and Sphagnum recurvum complex species (S. balticum and later S. fallax stem leaves were found). S. cuspidatum, which can grow in submerged conditions, occurs most abundantly ∼ 250–150 cal BP – falling at the end of the 18th and first half of the 19th century CE. This very wet phase is preceded by a high peak (25 %–70 %) of UOM ∼ 300–250 cal BP (1740–1650 CE), suggesting a disturbance around the second half of the 17th century CE and the first half of the 18th century. S. fallax, the dominant moss on the peatland today, has a confirmed continuous presence (stem leaves) since ∼190 cal BP (1750 CE) however, the exact proportion of this species among the identified recurvum complex species is uncertain. Rhododendron tomentosum macroremains (including seeds) are especially abundant in the samples dated to the early 19th century (depths 17.5 and 22.5 cm).
The phase is overall characterized by very rapid shifts in the testate amoebae-based water table fluctuations, although Sphagnum mosses show a stable wet period. Since the beginning of the phase, the abundance of H. papilio and A. flavum drops. In the samples with a high decomposition dated to ∼300–400 cal BP (1650–1570 CE) the count of tests is low. Around 200 cal BP (1740 CE) rapid fluctuations on the drier side begin. Nebela tincta, Alabasta militaris and G. discoides are abundant, referring to fluctuating water tables with dry periods (Lamentowicz et al., 2008a; Lamentowicz and Mitchell, 2005; Marcisz et al., 2015). Trigonopyxis arcula which appears abundantly in this zone is considered a drought-resistant species (Charman et al., 2007; Turner et al., 2014). The topmost analyzed sample (1.25 cm) is dominated by Corythion-Trinema type, which does not occur in deeper layers and results in a DWT of 33.3 cm. Other species that occur only in the uppermost peat samples include Aulacomnium palustre and Euglypha species (Figs. 5 and 6). The average DWT is 17.5 cm in this zone with a minimum of 7 and a maximum of 33.3 cm.
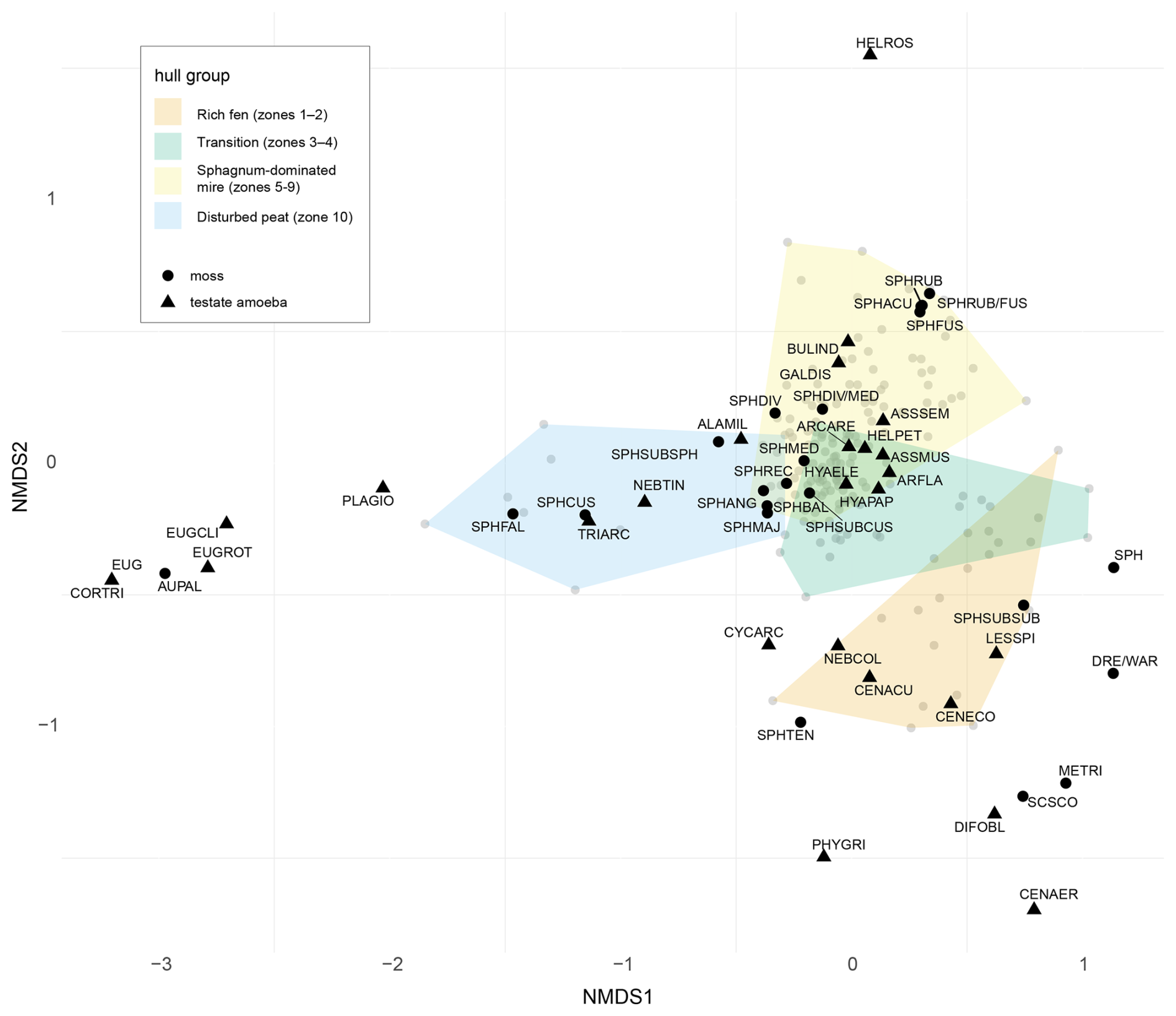
Figure 6Non-Metric Multidimensional Scaling (NMDS) ordination plot visualizing relationships between testate amoebae (black triangles) and bryophyte (black circles) species composition. Manually defined colored hulls represent different stages (samples in gray dots) in Linje mire development. Stress value: 0.1891065. Taxa abbreviations, Mosses: METRI = Meesia triquetra, SCSCO = Scorpidium scorpioides, AUPAL = Aulacomnium palustre, DRE/WAR = Drepanocladus/Warnstorfia-type, SPH = Sphagnum spp. (unspecified), SPHSUBSPH = Sphagnum subgenus Sphagnum, SPHDIV/MED = Sphagnum divinum/medium, SPHDIV = Sphagnum divinum, SPHMED = Sphagnum medium, SPHSECTACU = Sphagnum section Acutifolia, SPHRUB/FUS = Sphagnum rubellum/fuscum, SPHRUB = Sphagnum rubellum, SPHFUS = Sphagnum fuscum, SPHSUBCUS = Sphagnum subgenus Cuspidata, SPHCUS = Sphagnum cuspidatum, SPHMAJ = Sphagnum majus, SPHREC = Sphagnum recurvum complex, SPHBAL = Sphagnum balticum (stem leaves), SPHTEN = Sphagnum tenellum, SPHFAL = Sphagnum fallax (stem leaves), SPHANG = Sphagnum angustifolium (stem leaves), SPHSUBSUB = Sphagnum subgenus Subsecunda. Testate amoebae: ARFLA = Archerella flavum, ARCARE = Arcella arenaria, GALDIS = Galeripora discoides, ASSMUS = Assulina muscorum, ASSSEM = Assulina seminulum, BULIND = Bullinularia indica, CENACU = Centropyxis aculeata, CENAER = Centropyxis aerophila, CENECO = Centropyxis ecornis, CYCARC = Cyclopyxis arcelloides, DIFOBL = Difflugia oblonga, EUG = Euglypha sp., EUGCLI = Euglypha ciliata, EUGROT = Euglypha rotunda, HELPET = Heleopera petricola, HELROS = Heleopera rosea, HYAELE = Hyalosphenia elegans, HYAPAP = Hyalosphenia papilio, LESSPI = Lesquereusia sp., NEBCOL = Nebela colaris, ALAMIL = Alabasta militaris, NEBTIN = Nebela tincta, PHYGRI = Physochila griseola, PLAGIO = Plagiopyxis callida, TRIARC = Trigonopyxis arcula, CORTRI = Corythion-Trinema.
Multivariate analysis
NMDS ordination results (stress value: 0.1891065) (Fig. 6) illustrate four stages in Linje peatland development: (I) waterlogged rich fen (zones 1–2), (II) transitional phase from rich fen to poor fen (zones 3–4), (III) natural Sphagnum-dominated mire with various microforms (zones 5–9) and IV) disturbed peat layer (zone 10).
Species clustering on the positive side of NMDS1 and the negative end of NMDS2 (zones 1–2), represent the earliest stages of Linje peatland development, reflecting taxa preferring varying degrees of minerotrophic conditions (Gałka et al., 2013a; Hedenäs, 2003; Kalvaitienė and Jukonienė, 2025; Laine et al., 2018; Lamentowicz et al., 2007; Marcisz et al., 2020a; Stastney and Black, 2020). Among testate amoebae, these include C. ecornis, Centropyxis aculeata, Centropyxis aerophila, Difflugia oblonga, Lesquereusia sp. and C. arcelloides. Characteristic mosses are S. scorpioides, M. triquetra, Drepanocladus/Warnstorfia, Sphagnum subgenus Subsecunda and S. tenellum.
The following transitional period (zones 3–4) lacks clearly distinct species but overlaps with both the rich fen and Sphagnum-dominated assemblages, reflecting gradual ecological change.
The central cluster (around 0,0) comprises zones 5–9 and most species encountered in the Linje core. It corresponds to the resilient Sphagnum-dominated mire. Species here include the most common testate amoebae such as H. papilio, A. flavum and Assulina spp. and mosses S. divinum/medium and Sphagnum recurvum complex among others. Species characteristic of hummock microforms, such as Sphagnum mosses in the Acutifolia section, together with disturbance indicators (G. discoides) and hummock specialists (B. indica) (Lamentowicz et al., 2008a) occupy positions slightly toward the positive side of NMDS2.
Species found in the uppermost disturbed peat layer (zone 10) occupy the negative side of both axes. The testate amoebae in this side of the ordination are indicative of disturbances and suggest lower water tables. Mosses in this cluster include S. fallax, often linked to increased nutrient availability (Gąbka and Lamentowicz, 2008; Gałka and Lamentowicz, 2014; Limpens et al., 2003), and S. cuspidatum, a species of waterlogged conditions (De Vleeschouwer et al., 2012; Gałka et al., 2017b). This contradiction may reflect a seasonal disagreement between the proxies, caused by low-resolution sampling, that does not capture rapid changes. A. palustre and testate amoebae of Corythion-Trinema type and Euglypha species that occur only in the uppermost peat samples, form a separate group on the ordination plot.
Major shifts in the bryophyte assemblages correspond with established Holocene subdivisions, indicating wetter conditions during the Early and Late Holocene and drier conditions in the Middle Holocene (Wanner et al., 2011, 2015) (Figs. 3–5). Thus, the changes observed in these zones and their relationship to climatic and autogenic processes are analysed within the framework of the Early, Middle, and Late Holocene. Disturbance phases and shifts in bryophyte assemblages are considered in relation to periods of Rapid Climate Change (RCC) (Mayewski et al., 2004) (Fig. 7). However, because RCC intervals span several centuries to millennia and are characterized by fluctuating rather than uniform climate conditions, they cannot be directly equated with the hydrological changes and disturbances observed at Linje. Thus, RCC periods (and other global climatic events) are used here as a broader climatic framework within which to interpret the proxy-related changes at Linje.
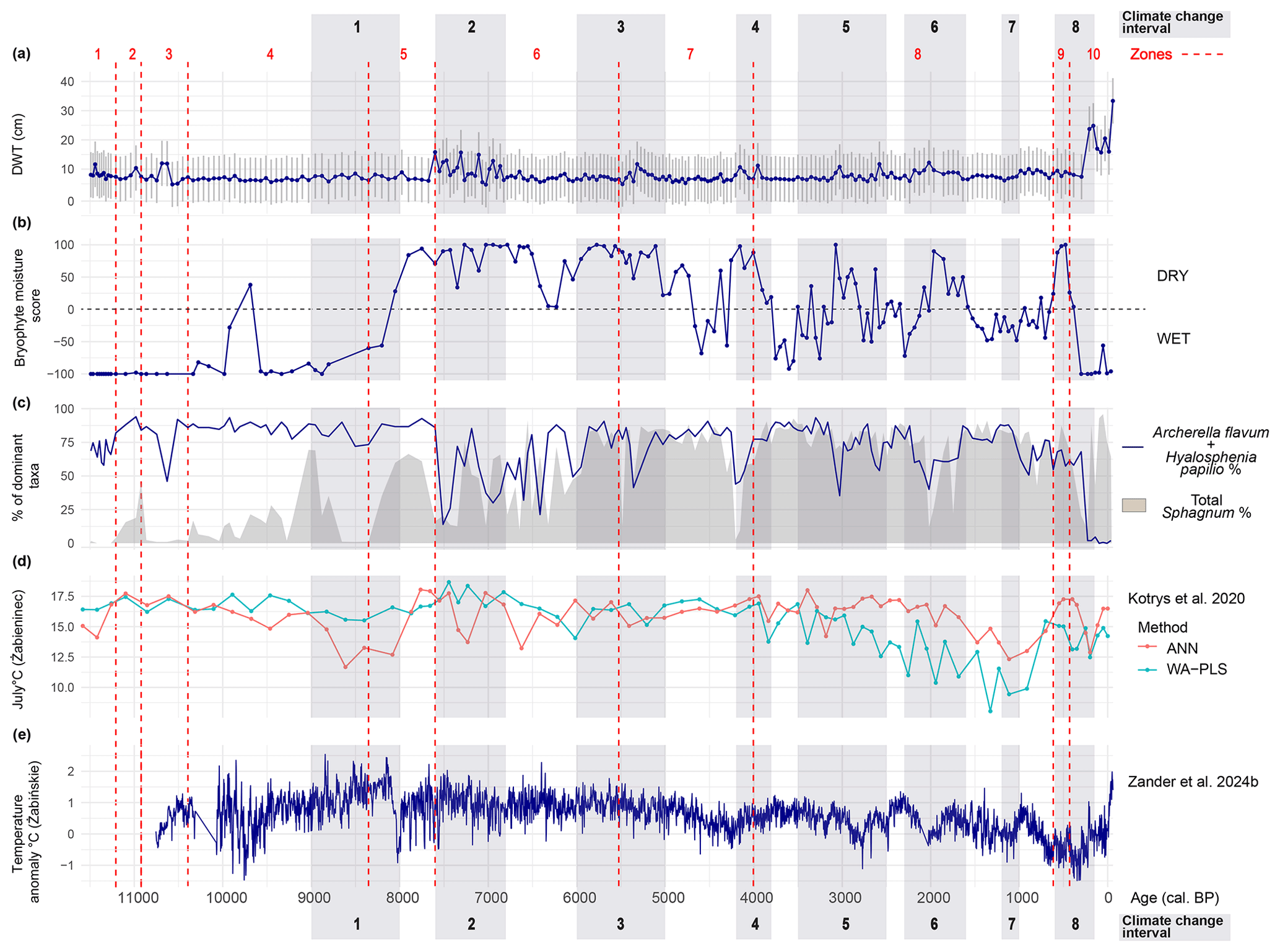
Figure 7Diagram showing local wetness changes and disturbance phases (climatic and anthropogenic) in Linje, and temperature variability in Poland during the Holocene (a) Testate-amoebae based depth to water table reconstruction (b) Bryophyte moisture indices (see method in chapter 3.4; Table A1) (c) Disturbances indicated by declines in Sphagnum or in dominant wet-indicator testate amoebae species (sum of A. flavum and H. papilio) (d) Reconstructed mean July air temperature (°C) from lake Żabieniec (central Poland) based on the chironomid training set using weighted averaging-partial least squares regression (WA-PLS) or an artificial neural network (ANN) transfer function (Kotrys et al., 2020) (e) Holocene spring-summer temperature anomaly reconstruction from lake Żabinìskie (northern Poland) based on ratios in varves (Zander et al., 2024b). Grey bars represent important climate change intervals recorded in Linje, RCC = period of rapid climate change (Mayewski et al., 2004): 1. RCC 9000–8000 cal BP, 2. Holocene Thermal Maximum in Linje (∼ 7600–6800 cal BP) 3. RCC 6000–5000 cal BP, 4. RCC 4200–3800 cal BP, 5. RCC 3500–2500 cal BP, 6. Roman Warm Period (2300–1600 cal BP) (Gauld et al., 2024), 7. RCC 1200–1000 cal BP (not recorded in Linje) 8. RCC 600–150 cal BP. Red dashed lines mark the bryophyte zones.
5.1 Early Holocene (Zones 1–5)
Peat formation in Linje mire began in the Early Holocene resulting from the short-term terrestrialization of a depression left by a dead ice block (Kloss, 2005; Kloss and Żurek, 2005). As the accumulation of peat started with brown mosses, there is a possibility that they initially grew on deposits overlying the buried ice block (Słowiński et al., 2015). Given that some dead ice blocks in northern Poland persisted until the Early Holocene if meltwater drainage was sufficient (Błaszkiewicz et al., 2015; Słowiński, 2010), it is possible that Linje depression was able to actively accumulate peat since the beginning of Holocene. In Linje, the sediments beneath the peat deposits consist of sand and gravel (Kloss and Żurek, 2005), suggesting that conditions for delayed melting may have been present. The dead ice should be marked by the presence of basal peat overlain by lacustrine sediments. These were not detected in the Linje profile. The layer of brown mosses suggests the development of a wet rich fen (with phases of short-term standing water) on the bottom of the site (Figs. 3 and 4). Therefore, this is not a typical situation as seen in other kettle-hole peatlands in Pomerania. Most kettle hole peatlands, in e.g. Tuchola Forest, possess brown moss peat under gyttja (Kowalewski, 2014; Lamentowicz, 2005). To sum up there are many pathways of so-called kettle-hole peatlands development (Brande et al., 1990; Succow and Joonsten, 2012; Timmermann, 1999, 2003, 2010; Timmermann and Succow, 2001) and there is a high uncertainty that dead ice played a role in the Linje basin formation. According to Żurek (2005) the dead ice block at Linje mire melted during the Allerød, as evidenced by basal brown moss peat dating. However, AMS 14C dates from our study (Fig. 2., Table 1), suggest that the onset of peat accumulation during the Allerød is questionable, as these previous dates (Kloss, 2005; Kloss and Żurek, 2005) were carried out from bulk peat so they might have been affected by reservoir effect i.e. revealing much older age, as brown mosses such as Drepanocladus sp. may also absorb old anions (Madeja and Latowski, 2008).
The transition from rich fen to poor fen in the centre of the peatland began ∼ 10 900 cal BP, as E. vaginatum began to form a suitable base for Sphagnum mosses to establish (Hughes and Dumayne-Peaty, 2002). A prerequisite for this process was the lowering/fluctuations of the water table during zone 3. The vegetation between ∼10 900 and ∼ 8350 cal BP is already suggesting a decrease in nutrients and increase in acidity. Peatland water table fluctuations in these early successional stages can most likely be attributed to autogenic peatland processes (Hughes et al., 2000).
The period between ∼ 9000–∼ 8000 cal BP is recognized as one of the periods of RCC and it is expressed as a cooling in the Northen Hemisphere (Mayewski et al., 2004). During this period pioneering oligotrophic Sphagnum communities became established in Linje, making the record more sensitive to climate fluctuations (Hughes and Barber, 2003). Between ∼8500 and ∼ 8350 cal BP samples from Linje show high decomposition, with a large proportion of UOM and an absence of Sphagnum. This suggests water table lowering, which aerated the acrotelm and accelerated decomposition. Concurrently, testate amoebae indicative of stable wet conditions show a decrease in abundance (Fig. 7c). This disturbance may be linked to the Early Holocene Thermal Maximum (Väliranta et al., 2015), that Zander et al. (2024a) have described as a period of maximum summer warmth in NE Poland between 8500 and 8100 cal BP (Fig. 7e). However, chironomid-based summer temperature reconstructions from Central Poland (170 km southeast of Linje) suggest slight cooling during this period (Kotrys et al., 2020) (Fig. 7d). Linje record does not resolve this apparent contradiction, owing to poor sample preservation during this period.
When Sphagnum recolonized after the disturbance (∼8200 cal BP), the species composition in Linje shifted from wetter microform species to those indicative of slightly drier conditions. This timing aligns with the well-documented 8.2 ka event, with strong evidence from multiple independent proxies across the Northern Hemisphere, including pollen-based reconstructions (Seppä et al., 2007; Veski et al., 2004), stable isotopes and geochemical analyses from lake sediments, including varves (Hammarlund, 2003; Fiłoc et al., 2017; Zander et al., 2024a), peat-based archives using testate amoebae and plant macrofossils (Gałka et al., 2014; Fletcher et al., 2024a), and stable peat isotopes (Daley et al., 2016). The hydrological response at the latitude of Linje and in Northern Europe is often associated with a drier climate during the 8.2 ka event (Magny et al., 2003, 2007), agreeing with the Linje record. At the same time the multi-proxy paleorecord of lake Suminko (NE Poland) shows a major lake-level rise between ∼ 8300 and ∼ 8100 cal BP (Pędziszewska et al., 2015). Overall, the period between ∼ 9000 and ∼ 8000 cal BP seems to be a time of hydrological instability in Linje peatland but the exact interpretation in relation to temperature, humidity or specific climate events remains uncertain because of the high degree of peat decomposition.
5.2 Middle Holocene (Zones 6–7)
The Holocene Thermal Maximum (HTM) was a period of elevated temperatures in the Northern Hemisphere, with noticeable effects in Northern Europe, including high-altitude tree migration, glacier retreat, and increased evapotranspiration (Davis et al., 2003, and references therein; Fletcher et al., 2024b; Kaufman et al., 2020; Wanner et al., 2015). Temperature reconstructions from Lake Żabińskie indicate the HTM in Poland began and peaked ∼ 8500 cal BP, with stable warm conditions lasting until 4700 cal BP (Fig. 7e) (Zander et al., 2024a, b). However, in Linje, bryophyte and testate amoebae records indicate the driest conditions between ∼7600 and ∼ 6800 cal BP (Figs. 3–5, and 7a and b). Around 7600 cal BP, high macrocharcoal percentages suggest increased local fire activity, while dominant testate amoebae and Sphagnum mosses experienced a simultaneous dramatic decline between ∼ 7600 and 7350 cal BP (Figs. 4 and 7c).
Pollen records from northern Finland also indicate the warmest summer temperatures and lowest precipitation between ∼ 7900 and ∼ 5700 cal BP (Seppä and Birks, 2001). Similarly, evidence of increased dryness appears in reduced peat formation rates in southern Finland (Korhola, 1995) and lowered water tables in southern Sweden ∼ 7500–7300 cal BP (Digerfeldt, 1988). Plant macrofossil and pollen records from a Polish lake (∼320 km northeast of Linje) suggest a corresponding temperature peak ∼ ∼7750 cal BP (Gałka et al., 2014). Chironomid-based summer temperature reconstructions from Lake Żabieniec further confirm a temperature peak either ∼7760–7450 or ∼7450–7230 cal BP, depending on the method used (Kotrys et al., 2020), aligning with Linje's testate amoebae and bryophyte records (Fig. 7d).
The onset of drier conditions in Linje aligns with several paleoclimate records that indicate Holocene temperature peaks following the 8.2 ka cooling event (Fletcher et al., 2024b; Wanner et al., 2015). However, this dry phase does not correspond with multiple regional climate reconstructions. For example, the period 7600–7250 cal BP has been identified as a cold phase based on pollen and aquatic invertebrate data from lakes in northeastern Poland (Fiłoc et al., 2017). The time interval between ∼ 8000 and 6800 cal BP is described to be favourable for peat initiation in Walton moss in England (Hughes et al., 2000) and in southern Finland (Korhola, 1995), indicating increased effective precipitation. These inconsistencies highlight how atmospheric and oceanic circulation shifts during this period caused regional climate anomalies and variations in the timing of the HTM (Fletcher et al., 2024b). Cartapanis et al. (2022), further note that the Northern Hemisphere HTM, as inferred from terrestrial proxies, lasted ∼ 4000 years (8–4 ka), making precise synchronization across regions difficult. Additionally, it cannot be ruled out that the dry shift observed in Linje reflects a localized autogenic succession.
Drier conditions persisted in Linje until ∼5500 cal BP, when a sudden peak in UOM occurs, followed by a gradual wet shift indicated by the bryophytes (Figs. 3, 4, and 7b and c). This coincides with “Mid-Holocene Cooling”, characterized by glacier advances and North-Atlantic ice-rafting events in many regions between ∼ 6000 and ∼ 5000 cal BP (Fletcher et al., 2024b; Kobashi et al., 2017; Mayewski et al., 2004; Wanner et al., 2015). A strong cooling signal has been recorded in varved lake sediments in eastern Poland ∼ 5500–5200 cal BP (Pędziszewska et al., 2015). Lake Gościąż (central Poland) also shows a higher water table between ∼ 6000 and ∼ 5200 cal BP (Pazdur et al., 1995). Similarly, in Gązwa bog, a period of increased humidity begins ∼ 5750 cal BP (Gałka and Lamentowicz, 2014). Multiple paleorecords across Europe also indicate a shift to wetter conditions during this time (see Table 4 in Hughes et al., 2000). Additionally, a wet shift (∼ 5800–4800 cal BP) has been recognised from subfossil Sphagnum lipid biomarkers in Ireland (Jordan et al., 2017). However, not all records show a uniform pattern of increased moisture. In contrast, Stążki peatland experienced a period of hydrological instability and drier conditions ∼5400 cal BP (Gałka et al., 2013b) and ∼ 5600 cal BP, lake-level lowering is observed in lake Linówek (northeastern Poland) (Gałka et al., 2014).
5.3 Late Holocene (Zones 8–10)
Between ∼4200 and ∼ 4150 cal BP, Linje experienced a brief disturbance marked by hydrological instability (Fig. 7a–c). This event likely corresponds to the 4.2 ka climatic event, which is often underrepresented in peat (Roland et al., 2014) but displays broad temporal variability in other paleo-proxies (Geirsdóttir et al., 2019; McKay et al., 2024). The 4.2 ka event is generally expressed as cooling or drought in the Northern Hemisphere (Yan and Liu, 2019). Hydrological instability or drought has also been recorded in other Polish peatlands coinciding with the 4.2 ka event (Gałka et al., 2013b; Lamentowicz et al., 2019b). A lake water decrease that resulted in a terrestrialization was also recorded ∼4150 cal BP in Rąbień mire (Central Poland) (Słowiński et al., 2016).
Between ∼3500 and ∼ 2500 cal BP Sphagnum and testate amoebae assemblages indicate a period of rapid hydrological fluctuations in Linje, agreeing with a RCC interval identified by Mayewski et al. (2004). Following this disturbance of the 4.2 ka event, the Sphagnum composition in Linje shifted to low-lawn and hollow species. Wetter conditions prevailed until ∼3250 cal BP, followed by a drier phase ∼3050 cal BP, when A. flavum and H. papilio abundance drops drastically (Fig. 7c). Moisture levels increased again towards the end of this period but remained variable. These fluctuations likely reflect the complex climatic dynamics between the Bronze Age Cold Epoch, the Bronze Age Optimum (characterized by drought and disturbance ∼ 3050 cal BP), and the Iron Age Cold Epoch, or the 2.8 ka event (Gauld et al., 2024; Kobashi et al., 2017). The latter is a widely documented wet shift in Europe (Barber et al., 2004; Mauquoy et al., 2008), including in Polish peat archives: in Kusowskie Bagno ∼2700 cal BP (Lamentowicz et al., 2015) and in Głęboczek ∼2800–2600 cal BP (Lamentowicz et al., 2019b). However, this pattern contrasts with the findings of Słowiński et al. (2016), who suggest that the 2.8 ka event in Europe followed a wet-to-dry gradient from west to east, with some Polish sites experiencing drier conditions. The hydrological response in Linje follows the pattern observed in Western Europe.
Another notable disturbance appears ∼ 2000 cal BP in plant and testate amoebae records (Fig. 7c), coinciding with the Roman Warm Period (Gauld et al., 2024), which is reflected in two Northern Ireland peatlands as drought episodes over an extended interval (Swindles et al., 2010). Similarly, lake sediment records from Northern Europe suggest an increase in dry conditions ∼ 2000 cal BP (Seppä et al., 2009). Pollen records shows that human impact in the vicinity of Linje during ∼2000 cal BP was low (Marcisz et al., 2015). However, human impact to the forest surrounding Linje is clearly visible in the pollen record between ∼3000 and ∼2500 cal BP (Szambelan et al., 2025), thus it is difficult to disentangle whether the observed disturbance is climate-driven or caused by human activity.
The period 1200–1000 cal BP is marked as a global RCC interval (Mayewski et al., 2004). Marcisz et al. (2015) have recorded a strong wet shift coinciding with this period, however in this work conditions are wet according to all proxies, but no major shifts or events are visible (Fig. 7a–c).
A relatively brief (around 200 years) dry period occurred in Linje between ∼600 and ∼ 450 cal BP characterised by changes in the Sphagnum composition in this section of the record. This shift can be associated with the climate fluctuations of the Little Ice Age (LIA) (Gauld et al., 2024; Marcisz et al., 2015, 2020a; Mauquoy et al., 2002). From a nearby coring location in Linje, a rapid shift to dry conditions (water table drop from 0 to 25 cm) in the testate amoebae record was found especially ∼ 550 cal BP (1390–1425 CE) (Marcisz et al., 2015), agreeing with the dry shift found in mosses in this work. Contradictory wet indicators, such as green algae, were also reported by Marcisz et al. (2015) and attributed to reduced evapotranspiration due to decreased forest cover and rapid climatic oscillations, causing temporary snowmelt excess. The dry phase in Linje coincides with hydrological instability recorded in peatlands from northern Poland, such as Słowińskie Błota (∼600 cal BP) (Lamentowicz et al., 2009b), ∼ 170 km northwest from Linje, Stążki (∼850–450 cal BP) (Lamentowicz et al., 2008b), located ∼ 140 km north from Linje and in Kusowskie Bagno (∼710–400 cal BP), a peatland ∼130 km northwest from Linje. These sites display high water table fluctuations, growth of E. vaginatum, and the disappearance of S. fuscum. In contrast, Linje exhibited persistent S. fuscum and S. rubellum growth, suggesting uninterrupted peat accumulation. Peatlands in central Poland might have had a different response as a multiproxy study from Żabieniec kettle hole contrastingly demonstrates a rapid wet shift at ∼600 cal BP (Lamentowicz et al., 2009a). In Saxnäs Mosse bog in Southern Sweden, the period of ∼675–360 cal BP records a cooling signal in the palynological data while the shifts in moss composition might indicate either a dry shift or an eutrophication signal (van der Linden and van Geel, 2006).
The drier period in Linje is interrupted ∼ 450 cal BP (1510 CE) when Sphagnum mosses indicate a sudden switch to wetter conditions. This coincides with the beginning of a prominent cooling during the LIA in Europe. The wet shift ∼ 450 cal BP is also observed in Stążki bog (Lamentowicz et al., 2008b), Żabieniec kettle hole (Lamentowicz et al., 2009a), Kusowskie Bagno (Lamentowicz et al., 2015), Finland (Väliranta et al., 2007) and Estonia (Sillasoo et al., 2007). However, the last 1000 years of climate records in Northern European peat show many alterations between dry and wet conditions that often do not align and are likely caused by differences in oceanic and continental climate (Marcisz et al., 2020b; Väliranta et al., 2007). The lack of spatial and temporal coherency during the LIA is also pointed out by Neukom et al. (2019) and possible climatic signals could be obscured by increasing human impact since early Medieval times resulting in deforestations and increasing openness affecting the hydrology of peatlands (Lamentowicz et al., 2009b). The early 16th-century wet phase may also reflect factors unrelated to climate. As Marcisz et al. (2015) point out, ∼430 cal BP (coinciding with the onset of the wet phase identified in this study) was marked by high fire activity in the Linje region, likely linked to military activities during the Polish-Teutonic war, suggesting that anthropogenic impact on the peatland was increasing.
Testate amoebae and bryophytes show contradicting information in the top 30 cm peat layer, which started ∼ 250 cal BP (1700 CE): testate amoebae show rapidly decreasing water tables and dry conditions whereas the Sphagnum taxa indicate wet, even waterlogged conditions (Figs. 3–6, and 7a–c). During these shifts, human impact in the vicinity of Linje was increasing, culminating with drainage in the 19th century CE, resulting in destabilized hydrology (Marcisz et al., 2015). Marcisz et al. (2014b) has found that the current testate amoebae communities in Linje differ significantly in spring and summer across all microforms. Thus, it is possible that the drought-resistant testate amoebae species recorded in the top layer reflect summer droughts that intensified after the drainage whereas plant macrofossils reflect the seasonal inundation and post-drainage peat subsidence. Current observations confirm that Linje's surface reacts sensitively to meteorological fluctuations due to drainage (Słowińska et al., 2022). The recent drying trend at Linje is consistent with patterns documented across many European peatlands (Swindles et al., 2019). The increased presence of S. fallax on the peatland might indicate a change in peatland chemistry, as S. fallax is more tolerant of nutrient inputs compared to the other Sphagnum mosses and the expansion of this species is connected to increased N deposition to nutrient-poor peatlands (Gąbka and Lamentowicz, 2008; Limpens et al., 2003). This change could be further investigated using geochemical analyses.
5.4 Site-specific considerations
Linje lies within a forest surrounded depression, that produces a distinctive microclimate compared to adjacent areas, with lower air temperatures and increased ground frost days (Słowińska et al., 2022). Although the forest cover surrounding the peatland has been dynamic (Marcisz et al., 2015; Szambelan et al., 2025), it is likely that during the formation of the peat layer in the depression, the site experienced a unique microclimate as well. Consequently, the peat profile's hydrological signals likely reflect both broader climate trends and local microclimatic conditions.
In addition to microclimate, peatland type should be considered while talking about climate sensitivity. Although we found in this study that the centre of Linje has been Sphagnum-dominated since ∼ 9050 cal BP, it should be considered that the peatland is not ombrotrophic as groundwater incursion influences its western side (Słowińska et al., 2010). This groundwater input cannot be overlooked, as it may have impacted the recorded hydrological shifts. However, other Sphagnum-dominated peatlands, beyond ombrotrophic bogs, have been recognized as valuable climate archives (Booth, 2010; Lamentowicz et al., 2008c). To know whether the shifts observed in Linje are climate-driven or products of internal peatland processes, it is essential to identify synchronous changes in other regional records (Hughes et al., 2000; Swindles et al., 2012), which is the case in Linje: after the peatland became Sphagnum-dominant, bryophyte species turnovers, the decreases in Sphagnum cover or dominant testate amoebae species closely coincide with major Holocene climatic events (Fig. 7).
5.5 Comparison to earlier work
One peat profile can provide detailed information on bog vegetation at a microform scale (Mauquoy and Yeloff, 2008). As observed by Słowińska et al. (2022), the microclimatic conditions within the microsites in Linje exhibit considerable variability. Ideally multiple cores from the same site could be used to reconstruct mesoscale ecohydrological changes through time (Barber et al., 1998; Mauquoy and Yeloff, 2008).
Among previous studies, the work by Marcisz et al. (2015), based on a nearby coring location within the same vegetation zone, is most directly comparable to this study. Covering the last 2000 years, their reconstruction of the upper 2 m of peat shows strong agreement with our results: showing a progression from wet to waterlogged conditions, followed by a brief dry phase, and culminating with hydrologically unstable conditions in the uppermost layer.
Earlier paleoecological studies at Linje offer valuable points of comparison, despite differences in methodology and data resolution. The stratigraphy outlined by Kloss and Żurek (2005) broadly agrees with the succession in the vegetation described in this work. From the same core, Kloss (2005) further described hydrological transformations in Linje based on a phytocoenotic moisture index, which presents a moisture gradient ranging from moderately wet to heavily waterlogged. This hydrological interpretation agrees with water table changes captured by testate amoebae in this study as it shows rather wet conditions throughout the Holocene. Moreover, a decrease in moisture ∼ 400 cal BP was also recorded in the work of Kloss (2005). However, a distinctly drier episode around the Middle Holocene is not recorded in this earlier paleohydrological work. The discrepancy could be attributed to the different resolution and methods employed for plant macrofossil analysis in Kloss's work (2005). Additionally, the analysed core in the work of Kloss (2005) is closer to the margin of the peatland (Kucharski and Kloss, 2005) located about 50 m southward from the coring site of this study.
A new full Holocene peat record from Sphagnum-dominated Linje peatland in northern Poland provides valuable insights into past wetness changes. Plant macrofossil and testate amoebae analysis reveal significant shifts in peatland hydrology and vegetation, driven by autogenic succession, climatic variability, and later anthropogenic impacts. Bryological species turnovers are recorded at ∼ 11 200, ∼ 10 350, ∼ 8200, ∼ 7500, ∼ 5500, ∼ 600, and ∼ 450 cal BP. Early Holocene changes appear to result from autogenic succession. Toward the end of the Early Holocene, peatland changes increasingly align with known periods of rapid climate change. Both plant macrofossil and testate amoebae proxies indicate drier conditions between ∼ 7600 and ∼ 6800 cal BP, coinciding with the Holocene Thermal Maximum. In the Late Holocene, the testate amoebae record indicates additional disturbances ∼ 3050, ∼ 2000, and ∼ 200 cal BP. While earlier episodes were short-lived, the most recent disturbance (∼200 cal BP) marks a permanent species turnover, reflecting a fundamental shift in peatland conditions driven by drainage.
For millennia, Linje peatland remained resilient to natural climate variability, maintaining peat accumulation despite repeated hydrological and vegetational shifts. However, following historical drainage, the system has not recovered, which highlights the limits of resilience once strong anthropogenic pressures are introduced. The Linje record confirms the complementary value of plant macrofossils, particularly bryophytes, and testate amoebae in reconstructing palaeohydrological conditions, and highlights the value of Sphagnum-dominated poor fens as climate archives.
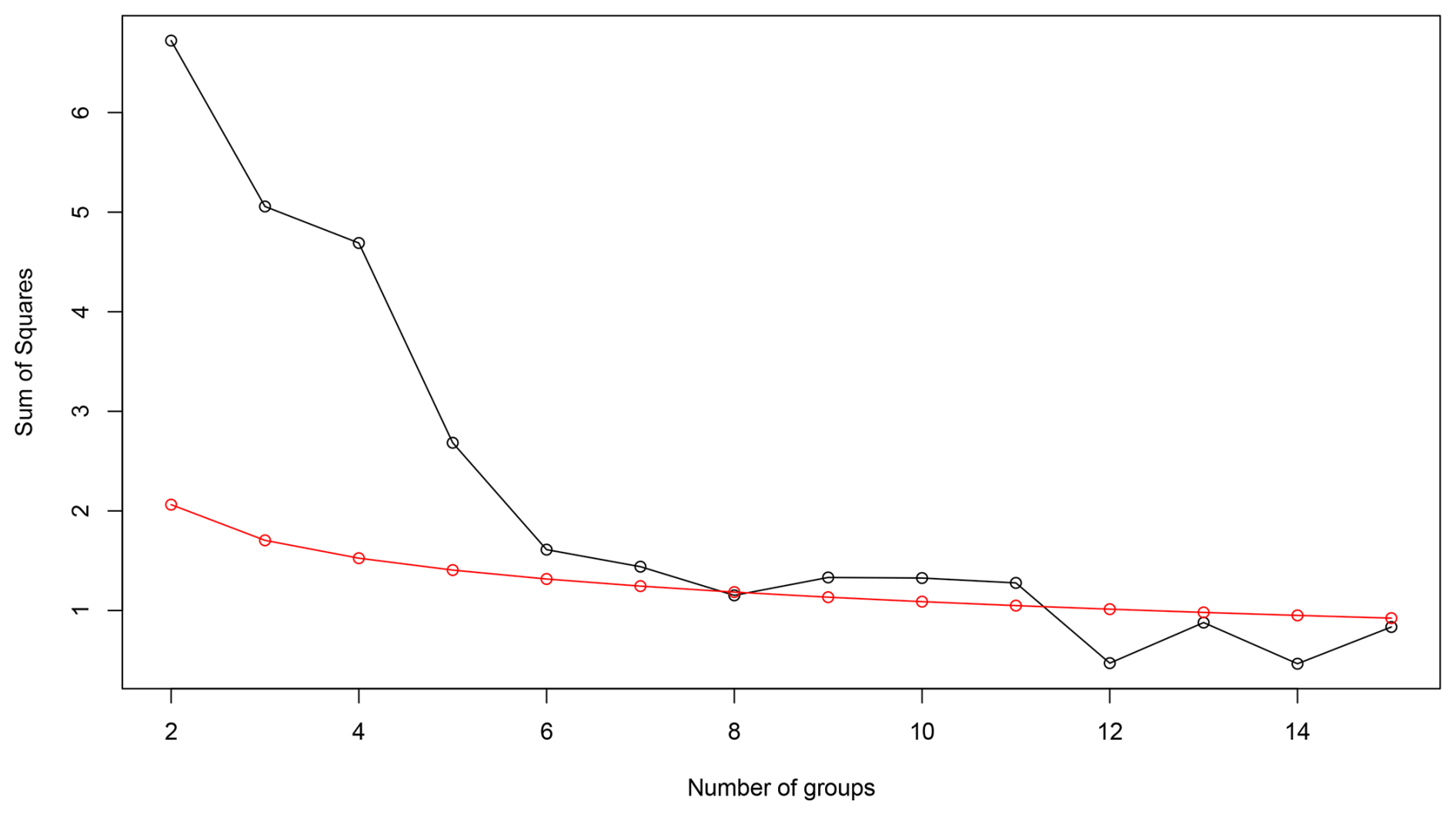
Figure A1Broken-stick model of bryophyte data from Linje peat core, showing the sum of squares (y axis) against the number of groups (x axis). The black line represents observed clustering results (CONISS) and the red line the expected broken-stick distribution. Clusters above the red line are considered significant. 11 clusters were initially identified, but cluster 8 was merged with cluster 7, giving a final solution of 10 clusters.
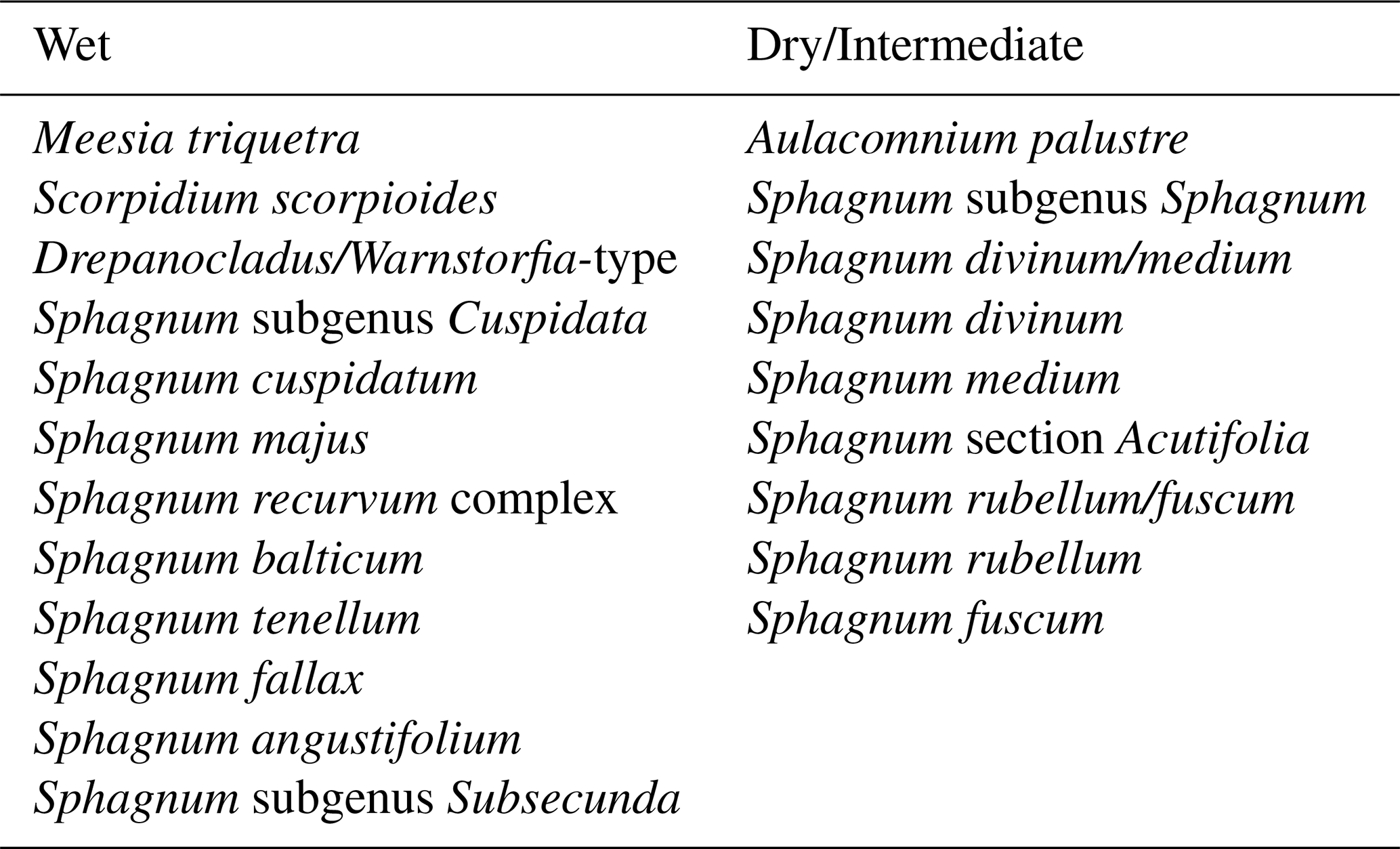
Table A1Assignment of bryophytes to wet or dry/intermediate groups (Hedenäs, 2003; Laine et al., 2018; Montagnes, 1990; Piilo et al., 2023). For the calculation of the bryophyte moisture curve, the percentages of wet-habitat species were transformed to negative values. Unidentified Sphagnum was excluded from the analysis due to uncertainties in habitat preference.
Plant macrofossil and testate amoebae data used in this study is stored in Mendeley Data repository: https://doi.org/10.17632/5f6565fdht.1 (Poolma et al., 2025).
EP prepared the manuscript, contributed to the statistical analysis, and visualization of the data, with help from KM and ML. EP also analysed plant macrofossils with guidance from LA and DM. ML organised the fieldwork plan and logistics. KM, LA, PK, MS, SV, and ML carried out the fieldwork. KM and PF analysed testate amoebae. KL and LA contributed to subsampling. KL and ML contributed to radiocarbon dating, for which KL aquired the funding. LA and ML aquired funding for the project. All authors contributed with their comments, and edits to the finalisation of the manuscript.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We thank Triin Reitalu, Fabian EZ Ercan, Ansis Blaus, Dominika Łuców and Sambor Czerwiński for the help during the fieldwork.
This research was supported by ESF project PRG1993, the Doctoral School of Tallinn University of Technology and the Ministry of Education and Research Centres of Excellence grant TK215. The study was supported by the National Science Centre, Poland, grants no 2021/41/B/ST10/00060, 2020/39/D/ST10/00641, 2019/03/X/ST10/00488 and Funding For A Joint Polish – Estonian Project Under The Agreement On Scientific Cooperation Between The Polish Academy Of Sciences And Estonian Academy Of Sciences. The study also benefited from an Erasmus Plus grant no. KA131 (Adam Mickiewicz University, 2019).
This paper was edited by Stephen Obrochta and reviewed by three anonymous referees.
Adobe Inc.: Adobe Illustrator CC (version 23.0), Adobe Systems Incorporated, San Jose, California, USA [software], 2019.
Amesbury, M. J., Swindles, G. T., Bobrov, A., Charman, D. J., Holden, J., Lamentowicz, M., Mallon, G., Mazei, Y., Mitchell, E. A. D., Payne, R. J., Roland, T. P., Turner, T. E., and Warner, B. G.: Development of a new pan-European testate amoeba transfer function for reconstructing peatland palaeohydrology, Quaternary Science Reviews, 152, 132–151, https://doi.org/10.1016/j.quascirev.2016.09.024, 2016.
Banaś, K., Ronowski, R., and Marciniak, P.: Effects of Environmental Conditions on the Individual Architectures and Photosynthetic Performances of Three Species in Drosera, International Journal of Molecular Sciences, 24, 9823, https://doi.org/10.3390/ijms24129823, 2023.
Barber, K. E., Chambers, F. M., Maddy, D., Stoneman, R., and Brew, J. S.: A sensitive high-resolution record of late Holocene climatic change from a raised bog in northern England, The Holocene, 4, 198–205, https://doi.org/10.1177/095968369400400209, 1994.
Barber, K. E., Dumayne-Peaty, L., Hughes, P., Mauquoy, D., and Scaife, R.: Replicability and variability of the recent macrofossil and proxy-climate record from raised bogs: field stratigraphy and macrofossil data from Bolton Fell Moss and Walton Moss, Cumbria, England, J. Quaternary Sci., 13, 515–528, https://doi.org/10.1002/(SICI)1099-1417(1998110)13:6<515::AID-JQS393>3.0.CO;2-S, 1998.
Barber, K. E., Chambers, F., and Maddy, D.: Late Holocene climatic history of northern Germany and Denmark: peat macrofossil investigations at Dosenmoor, Schleswig-Holstein, and Svanemose, Jutland, Boreas, 33, 132–144, https://doi.org/10.1080/03009480410001082, 2004.
Bennett, K. D.: Determination of the number of zones in a biostratigraphical sequence, New Phytologist, 132, 155–170, https://doi.org/10.1111/j.1469-8137.1996.tb04521.x, 1996.
Birks, H. J. B. and Seppä, H.: Late-Quaternary palaeoclimatic research in Fennoscandia – A historical review, Boreas, 39, 655–673, https://doi.org/10.1111/j.1502-3885.2010.00160.x, 2010.
Blackford, J.: Palaeoclimatic records from peat bogs, Trends in Ecology and Evolution, 15, 193–198, https://doi.org/10.1016/S0169-5347(00)01826-7, 2000.
Błaszkiewicz, M., Piotrowski, J. A., Brauer, A., Gierszewski, P., Kordowski, J., Kramkowski, M., Lamparski, P., Lorenz, S., Noryśkiewicz, A. M., Ott, F., Słowiński, M., and Tyszkowski, S.: Climatic and morphological controls on diachronous postglacial lake and river valley evolution in the area of Last Glaciation, northern Poland, Quaternary Science Reviews, 109, 13–27, https://doi.org/10.1016/j.quascirev.2014.11.023, 2015.
Bonk, A., Müller, D., Ramisch, A., Kramkowski, M. A., Noryśkiewicz, A. M., Sekudewicz, I., Gąsiorowski, M., Luberda-Durnaś, K., Słowiński, M., Schwab, M., Tjallingii, R., Brauer, A., and Błaszkiewicz, M.: Varve microfacies and chronology from a new sediment record of Lake Gościąż (Poland), Quaternary Science Reviews, 251, 106715, https://doi.org/10.1016/j.quascirev.2020.106715, 2021.
Booth, R. K.: Testing the climate sensitivity of peat-based paleoclimate reconstructions in mid-continental North America, Quaternary Science Reviews, 29, 720–731, https://doi.org/10.1016/j.quascirev.2009.11.018, 2010.
Booth, R. K., Lamentowicz, M., and Charman, D. J.: Preparation and analysis of testate amoebae in peatland palaeoenvironmental studies, Mires and Peat, 7, 1–7, 2010.
Brande, A., Hoelzmann, P., and Klawitter, J.: Genese und Paläoökologie eines brandenburgischen Kesselmoores, Telma, 20, 27–54, 1990.
Bronk Ramsey, C.: Radiocarbon calibration and analysis of stratigraphy: the OxCal program, Radiocarbon, 37, 425–430, https://doi.org/10.1017/S0033822200030903, 1995.
Bronk Ramsey, C.: OxCal program v 4.0, University of Oxford [code], https://c14.arch.ox.ac.uk/oxcal.html (last access: 4 February 2024), 2006.
Bronk Ramsey, C.: Deposition models for chronological records, Quaternary Science Reviews, 27, 42–60, https://doi.org/10.1016/j.quascirev.2007.01.019, 2008.
Bronk Ramsey, C. and Lee, S.: Recent and planned developments of the program OxCal, Radiocarbon, 55, 720–730, https://doi.org/10.1017/S0033822200057878, 2013.
Buttler, A., Bragazza, L., Laggoun-Défarge, F., Gogo, S., Toussaint, M., Lamentowicz, M., Chojnicki, B. H., Słowiński, M., Słowińska, S., Zielińska, M., Reczuga, M., Barabach, J., Marcisz, K., Lamentowicz, Ł., Harenda, K., Lapshina, E., Gilbert, D., Schlaepfer, R., and Jassey, V. E. J.: Ericoid shrub encroachment shifts aboveground–belowground linkages in three peatlands across Europe and Western Siberia, Global Change Biology, 29, 6772–6793, https://doi.org/10.1111/gcb.16904, 2023.
Cappers, R. T. J., Bekker, R. M., and Jans, J. E. A.: Digitale zadenatlas van Nederland: Digital seed atlas of the Netherlands, 2nd ed., Barkhuis and Groningen University Library, Groningen, 502 pp., ISBN 978-90-77922-95-8, 2012.
Cartapanis, O., Jonkers, L., Moffa-Sanchez, P., Jaccard, S. L., and De Vernal, A.: Complex spatio-temporal structure of the Holocene Thermal Maximum, Nat. Commun., 13, 5662, https://doi.org/10.1038/s41467-022-33362-1, 2022.
Chambers, F. M. and Charman, D. J.: Holocene environmental change: contributions from the peatland archive, The Holocene, 14, 1–6, https://doi.org/10.1191/0959683604hl684ed, 2004.
Chambers, F. M., Booth, R. K., De Vleeschouwer, F., Lamentowicz, M., Le Roux, G., Mauquoy, D., Nichols, J. E., and van Geel, B.: Development and refinement of proxy-climate indicators from peats, Quaternary International, 268, 21–33, https://doi.org/10.1016/j.quaint.2011.04.039, 2012.
Charman, D. J., Blundell, A., Chiverrell, R. C., Hendon, D., and Langdon, P. G.: Compilation of non-annually resolved Holocene proxy climate records: stacked Holocene peatland palaeo-water table reconstructions from northern Britain, Quaternary Science Reviews, 25, 336–350, https://doi.org/10.1016/j.quascirev.2005.05.005, 2006.
Charman, D. J., Blundell, A., and ACCROTELM Members: A new European testate amoebae transfer function for palaeohydrological reconstruction on ombrotrophic peatlands, J. Quaternary Science, 22, 209–221, https://doi.org/10.1002/jqs.1026, 2007.
Charman, D. J., Barber, K. E., Blaauw, M., Langdon, P. G., Mauquoy, D., Daley, T. J., Hughes, P. D. M., and Karofeld, E.: Climate drivers for peatland palaeoclimate records, Quaternary Science Reviews, 28, 1811–1819, https://doi.org/10.1016/j.quascirev.2009.05.013, 2009.
Clark, J. S.: Particle Motion and the Theory of Charcoal Analysis: Source Area, Transport, Deposition, and Sampling, Quat. Res., 30, 67–80, https://doi.org/10.1016/0033-5894(88)90088-9, 1988.
Clarke, K. J.: Guide to Identification of Soil Protozoa – Testate Amoebae., Freshwater Biological Association, Ambleside, U.K., ISBN 0-900386-69-X, 2003.
Clymo, R. S.: The Limits to Peat Bog Growth, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 303, 605–654, 1984.
Daley, T. J., Barber, K. E., Hughes, P. D. M., Loader, N. J., Leuenberger, M., and Street-Perrott, F. A.: The 8.2 ka BP event in north-eastern North America: first combined oxygen and hydrogen isotopic data from peat in Newfoundland, J. Quaternary Science, 31, 416–425, https://doi.org/10.1002/jqs.2870, 2016.
Davies, M. A., McLaughlin, J. W., Packalen, M. S., and Finkelstein, S. A.: Using Water Table Depths Inferred From Testate Amoebae to Estimate Holocene Methane Emissions From the Hudson Bay Lowlands, Canada, JGR Biogeosciences, 126, e2020JG005969, https://doi.org/10.1029/2020JG005969, 2021.
Davis, B. A. S., Brewer, S., Stevenson, A. C., and Guiot, J.: The temperature of Europe during the Holocene reconstructed from pollen data, Quaternary Science Reviews, 22, 1701–1716, https://doi.org/10.1016/S0277-3791(03)00173-2, 2003.
De Vleeschouwer, F., Pazdur, A., Luthers, C., Streel, M., Mauquoy, D., Wastiaux, C., Le Roux, G., Moschen, R., Blaauw, M., Pawlyta, J., Sikorski, J., and Piotrowska, N.: A millennial record of environmental change in peat deposits from the Misten bog (East Belgium), Quaternary International, 268, 44–57, https://doi.org/10.1016/j.quaint.2011.12.010, 2012.
Digerfeldt, G.: Reconstruction and regional correlation of Holocene lake-level fluctuations in Lake Bysjön, South Sweden, Boreas, 17, 165–182, https://doi.org/10.1111/j.1502-3885.1988.tb00544.x, 1988.
Drzymulska, D.: Mosses recognized as glacial relicts from their postglacial distribution in Poland, Veget. Hist. Archaeobot., 33, 657–669, https://doi.org/10.1007/s00334-023-00983-5, 2024.
Duffy, A. M., Aguero, B., Stenøien, H. K., Flatberg, K. I., Ignatov, M. S., Hassel, K., and Shaw, A. J.: Phylogenetic structure in the Sphagnum recurvum complex (Bryophyta) in relation to taxonomy and geography, Appl. Plant. Sci., 107, 1283–1295, https://doi.org/10.1002/ajb2.1525, 2020.
Fiłoc, M., Kupryjanowicz, M., Szeroczyńska, K., Suchora, M., and Rzodkiewicz, M.: Environmental changes related to the 8.2 ka event and other climate fluctuations during the middle Holocene: Evidence from two dystrophic lakes in NE Poland, The Holocene, 27, 1550–1566, https://doi.org/10.1177/0959683617702233, 2017.
Finlayson, C. M. and Spiers, A. G. (Eds.): Global review of wetland resources and priorities for wetland inventory, Supervising Scientist, Canberra, 520 pp., ISBN 978-0-642-24347-8, 1999.
Fletcher, W. J., Sánchez Goñi, M. F., Naughton, F., and Seppä, H.: Greenlandian Stage (Early Holocene, 11.7–8.2 ka), in: European Glacial Landscapes, Elsevier, 73–87, https://doi.org/10.1016/B978-0-323-99712-6.00029-5, 2024a.
Fletcher, W. J., Sánchez Goñi, M. F., Naughton, F., and Seppä, H.: Northgrippian Stage (Middle Holocene, 8.2–4.2 ka), in: European Glacial Landscapes, Elsevier, 89–104, https://doi.org/10.1016/B978-0-323-99712-6.00027-1, 2024b.
Gąbka, M. and Lamentowicz, M.: Vegetation-Environment Relationships in Peatlands Dominated by Sphagnum fallax in Western Poland, Folia Geobot., 43, 413–429, https://doi.org/10.1007/s12224-008-9023-8, 2008.
Gałka, M. and Lamentowicz, M.: Sphagnum succession in a Baltic bog in central-eastern Europe over the last 6200 years and paleoecology of Sphagnum contortum, The Bryologist, 117, 22–36, https://doi.org/10.1639/0007-2745-117.1.022, 2014.
Gałka, M., Lamentowicz, Ł., and Lamentowicz, M.: Palaeoecology of Sphagnum obtusum in NE Poland, The Bryologist, 116, 238–247, https://doi.org/10.1639/0007-2745-116.3.238, 2013a.
Gałka, M., Miotk-Szpiganowicz, G., Goslar, T., Jęśko, M., van der Knaap, W. O., and Lamentowicz, M.: Palaeohydrology, fires and vegetation succession in the southern Baltic during the last 7500 years reconstructed from a raised bog based on multi-proxy data, Palaeogeography, Palaeoclimatology, Palaeoecology, 370, 209–221, https://doi.org/10.1016/j.palaeo.2012.12.011, 2013b.
Gałka, M., Tobolski, K., Zawisza, E., and Goslar, T.: Postglacial history of vegetation, human activity and lake-level changes at Jezioro Linówek in northeast Poland, based on multi-proxy data, Veget. Hist. Archaeobot., 23, 123–152, https://doi.org/10.1007/s00334-013-0401-7, 2014.
Gałka, M., Tobolski, K., Górska, A., and Lamentowicz, M.: Resilience of plant and testate amoeba communities after climatic and anthropogenic disturbances in a Baltic bog in Northern Poland: Implications for ecological restoration, The Holocene, 27, 130–141, https://doi.org/10.1177/0959683616652704, 2017a.
Gałka, M., Tobolski, K., Lamentowicz, Ł., Ersek, V., Jassey, V. E. J., van der Knaap, W. O., and Lamentowicz, M.: Unveiling exceptional Baltic bog ecohydrology, autogenic succession and climate change during the last 2000 years in CE Europe using replicate cores, multi-proxy data and functional traits of testate amoebae, Quaternary Science Reviews, 156, 90–106, https://doi.org/10.1016/j.quascirev.2016.11.034, 2017b.
Gauld, J., Fletcher, W. J., Sánchez Goñi, M. F., Naughton, F., and Seppä, H.: Meghalayan Stage (Late Holocene, 4.2 ka–present), in: European Glacial Landscapes, Elsevier, 105–126, https://doi.org/10.1016/B978-0-323-99712-6.00028-3, 2024.
GBIF Secretariat: GBIF Backbone Taxonomy, https://doi.org/10.15468/39OMEI, 2023.
Geirsdóttir, Á., Miller, G. H., Andrews, J. T., Harning, D. J., Anderson, L. S., Florian, C., Larsen, D. J., and Thordarson, T.: The onset of neoglaciation in Iceland and the 4.2 ka event, Clim. Past, 15, 25–40, https://doi.org/10.5194/cp-15-25-2019, 2019.
Gong, J., Roulet, N., Frolking, S., Peltola, H., Laine, A. M., Kokkonen, N., and Tuittila, E.-S.: Modelling the habitat preference of two key Sphagnum species in a poor fen as controlled by capitulum water content, Biogeosciences, 17, 5693–5719, https://doi.org/10.5194/bg-17-5693-2020, 2020.
Granlund, L., Vesakoski, V., Sallinen, A., Kolari, T. H. M., Wolff, F., and Tahvanainen, T.: Recent Lateral Expansion of Sphagnum Bogs Over Central Fen Areas of Boreal Aapa Mire Complexes, Ecosystems, 25, 1455–1475, https://doi.org/10.1007/s10021-021-00726-5, 2022.
Grimm, E. C.: CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares, Computers and Geosciences, 13, 13–35, https://doi.org/10.1016/0098-3004(87)90022-7, 1987.
Grimm, E. C.: Tilia software (version 3.0.1), Illinois State Museum, Springfield, IL, USA [code], https://www.neotomadb.org/apps/tilia (last access: 27 October 2025), 2011.
Grosse-Brauckmann, G.: Über pflanzliche Makrofossilien mitteleuropäischer Torfe, https://doi.org/10.23689/FIDGEO-5276, 1972.
Hájková, P., Horsák, M., Hájek, M., Jankovská, V., Jamrichová, E., and Moutelíková, J.: Using multi-proxy palaeoecology to test a relict status of refugial populations of calcareous-fen species in the Western Carpathians, The Holocene, 25, 702–715, https://doi.org/10.1177/0959683614566251, 2015.
Halsey, L. A., Vitt, D. H., and Gignac, L. D.: Sphagnum-dominated Peatlands in North America Since the Last Glacial Maximum: Their Occurrence and Extent, The Bryologist, 103, 334–352, https://doi.org/10.1639/0007-2745(2000)103[0334:SDPINA]2.0.CO;2, 2000.
Hammarlund, D.: Rapid hydrological changes during the Holocene revealed by stable isotope records of lacustrine carbonates from Lake Igelsjön, southern Sweden, Quaternary Science Reviews, 22, 353–370, https://doi.org/10.1016/S0277-3791(02)00091-4, 2003.
Hassel, K., Kyrkjeeide, M. O., Yousefi, N., Prestø, T., Stenøien, H. K., Shaw, J. A., and Flatberg, K. I.: Sphagnum divinum (sp. nov.) and S. medium Limpr. and their relationship to S. magellanicum Brid., Journal of Bryology, 40, 197–222, https://doi.org/10.1080/03736687.2018.1474424, 2018.
Hedenäs, L.: The European species of the Calliergon-Scorpidium-Drepanocladus complex, including some related or similar species, Meylania, 28, 1–116, 2003.
Holmquist, J. R., Booth, R. K., and MacDonald, G. M.: Boreal peatland water table depth and carbon accumulation during the Holocene thermal maximum, Roman Warm Period, and Medieval Climate Anomaly, Palaeogeography, Palaeoclimatology, Palaeoecology, 444, 15–27, https://doi.org/10.1016/j.palaeo.2015.11.035, 2016.
Hughes, P. D. M. and Barber, K. E.: Mire development across the fen–bog transition on the Teifi floodplain at Tregaron Bog, Ceredigion, Wales, and a comparison with 13 other raised bogs, Journal of Ecology, 91, 253–264, 2003.
Hughes, P. D. M. and Dumayne-Peaty, L.: Testing theories of mire development using multiple successions at Crymlyn Bog, West Glamorgan, South Wales, UK, Journal of Ecology, 90, 456–471, https://doi.org/10.1046/j.1365-2745.2002.00677.x, 2002.
Hughes, P. D. M., Mauquoy, D., Barber, K. E., and Langdon, P. G.: Mire-development pathways and palaeoclimatic records from a full Holocene peat archive at Walton Moss, Cumbria, England, The Holocene, 10, 465–479, https://doi.org/10.1191/095968300675142023, 2000.
Hughes, P. D. M., Blundell, A., Charman, D. J., Bartlett, S., Daniell, J. R. G., Wojatschke, A., and Chambers, F. M.: An 8500cal. year multi-proxy climate record from a bog in eastern Newfoundland: contributions of meltwater discharge and solar forcing, Quaternary Science Reviews, 25, 1208–1227, https://doi.org/10.1016/j.quascirev.2005.11.001, 2006.
Jassey, V. E. J., Reczuga, M. K., Zielińska, M., Słowińska, S., Robroek, B. J. M., Mariotte, P., Seppey, C. V. W., Lara, E., Barabach, J., Słowiński, M., Bragazza, L., Chojnicki, B. H., Lamentowicz, M., Mitchell, E. A. D., and Buttler, A.: Tipping point in plant–fungal interactions under severe drought causes abrupt rise in peatland ecosystem respiration, Global Change Biology, 24, 972–986, https://doi.org/10.1111/gcb.13928, 2018.
Jaszczuk, I., Jabłońska, E., Kozub, Ł., Tanneberger, F., Aggenbach, C., Seeber, E., van Diggelen, R., Kreyling, J., Silvennoinen, H. M., and Kotowski, W.: Peat formation potential of temperate fens increases with hydrological stability, Science of The Total Environment, 947, 174617, https://doi.org/10.1016/j.scitotenv.2024.174617, 2024.
Jordan, S. F., Murphy, B. T., O'Reilly, S. S., Doyle, K. P., Williams, M. D., Grey, A., Lee, S., McCaul, M. V., and Kelleher, B. P.: Mid-Holocene climate change and landscape formation in Ireland: Evidence from a geochemical investigation of a coastal peat bog, Organic Geochemistry, 109, 67–76, https://doi.org/10.1016/j.orggeochem.2017.02.004, 2017.
Juggins, S.: C2 Version 1.5 User guide. Software for ecological and palaeoecological data analysis and visualisation, Newcastle University, Newcastle upon Tyne, UK [code], https://www.academia.edu/download/84693763/C2.pdf (last access: 27 October 2025), 2007.
Juggins, S.: rioja: Analysis of Quaternary Science Data, Comprehensive R Archive Network (CRAN) [code], https://doi.org/10.32614/CRAN.package.rioja, 2009.
Kalvaitienė, M. and Jukonienė, I.: The Ecological Drivers of the Mosaic Structure of Bryophyte and Vascular Plant Cover in the Rich Fens of Lithuania, Plants, 14, 2662, https://doi.org/10.3390/plants14172662, 2025.
Karpińska-Kołaczek, M., Kołaczek, P., Marcisz, K., Gałka, M., Kajukało-Drygalska, K., Mauquoy, D., and Lamentowicz, M.: Kettle-hole peatlands as carbon hot spots: Unveiling controls of carbon accumulation rates during the last two millennia, Catena, 237, 107764, https://doi.org/10.1016/j.catena.2023.107764, 2024.
Katz, N. J., Katz, S. V., and Skobeyeva, E.:. Atlas of plant remains in peat., Nedra, Moscow, 1005, 1977 (in Russian).
Kaufman, D., McKay, N., Routson, C., Erb, M., Dätwyler, C., Sommer, P. S., Heiri, O., and Davis, B.: Holocene global mean surface temperature, a multi-method reconstruction approach, Sci. Data, 7, 201, https://doi.org/10.1038/s41597-020-0530-7, 2020.
Kloss, M.: Identification of subfossil plant communities and palaeohydrological changes in raised mire development, Monographiae Botanicae, 94, 81–116, 2005.
Kloss, M. and Żurek, S.: Geology of raised mire deposits, Monographiae Botanicae, 94, 65–80, 2005.
Kobashi, T., Menviel, L., Jeltsch-Thömmes, A., Vinther, B. M., Box, J. E., Muscheler, R., Nakaegawa, T., Pfister, P. L., Döring, M., Leuenberger, M., Wanner, H., and Ohmura, A.: Volcanic influence on centennial to millennial Holocene Greenland temperature change, Sci. Rep., 7, 1441, https://doi.org/10.1038/s41598-017-01451-7, 2017.
Kokkonen, N. A. K., Laine, A. M., Laine, J., Vasander, H., Kurki, K., Gong, J., and Tuittila, E.: Responses of peatland vegetation to 15 year water level drawdown as mediated by fertility level, J. Vegetation Science, 30, 1206–1216, https://doi.org/10.1111/jvs.12794, 2019.
Korhola, A.: Holocene climatic variations in southern Finland reconstructed from peat-initiation data, The Holocene, 5, 43–57, https://doi.org/10.1177/095968369500500106, 1995.
Kotrys, B., Płóciennik, M., Sydor, P., and Brooks, S. J.: Expanding the Swiss-Norwegian chironomid training set with Polish data, Boreas, 49, 89–107, https://doi.org/10.1111/bor.12406, 2020.
Kowalewski, G.: Alogeniczne i autogeniczne składowe zarastania jezior: hipoteza wahań poziomu wody, Polskie Towarzystwo Limnologiczne, Bogucki Wydawnictwo Naukowe, Poznań, Poland, 2014.
Kucharski, L. and Kloss, M.: Contemporary vegetation of selected raised mires and its preservation, Monographiae Botanicae, 94, 37–64, 2005.
Kuuri-Riutta, O., Väliranta, M., and Tuittila, E.-S.: Literature review on testate amoebae as environmental indicators and as a functional part of the microbial community in northern peatlands, Mires Peat, 28, 1–16, ISSN 1819-754X, 2022.
Laine, J., Flatberg, K. I., Harju, P., Timonen, T., Minkkinen, K., Laine, A., Tuittila, E.-S., and Vasander, H.: Sphagnum mosses: the stars of European mires, Department of Forest Sciences, University of Helsinki, Helsinki, 326 pp., ISBN 951-51-3143-X, 2018.
Lamentowicz, Ł., Lamentowicz, M., and Gąbka, M.: Testate amoebae ecology and a local transfer function from a peatland in western Poland, Wetlands, 28, 164–175, https://doi.org/10.1672/07-92.1, 2008a.
Lamentowicz, M.: Geneza torfowisk naturalnych i seminaturalnych w Nadleśnictwie Tuchola (Origin and development of natural and seminatural peatlands in Tuchola Forest Inspectorate), Bogucki Wydawnictwo Naukowe, ISBN 83-60247-14-5, 2005.
Lamentowicz, M. and Mitchell, E. A. D.: The Ecology of Testate Amoebae (Protists) in Sphagnum in North-western Poland in Relation to Peatland Ecology, Microb. Ecol., 50, 48–63, https://doi.org/10.1007/s00248-004-0105-8, 2005.
Lamentowicz, M., Tobolski, K., and Mitchell, E. A. D.: Palaeoecological evidence for anthropogenic acidification of a kettle-hole peatland in northern Poland, The Holocene, 17, 1185–1196, https://doi.org/10.1177/0959683607085123, 2007.
Lamentowicz, M., Cedro, A., Gałka, M., Goslar, T., Miotk-Szpiganowicz, G., Mitchell, E. A. D., and Pawlyta, J.: Last millennium palaeoenvironmental changes from a Baltic bog (Poland) inferred from stable isotopes, pollen, plant macrofossils and testate amoebae, Palaeogeography, Palaeoclimatology, Palaeoecology, 265, 93–106, https://doi.org/10.1016/j.palaeo.2008.04.023, 2008b.
Lamentowicz, M., Obremska, M., and Mitchell, E. A. D.: Autogenic succession, land-use change, and climatic influences on the Holocene development of a kettle-hole mire in Northern Poland, Review of Palaeobotany and Palynology, 151, 21–40, https://doi.org/10.1016/j.revpalbo.2008.01.009, 2008c.
Lamentowicz, M., Balwierz, Z., Forysiak, J., Płóciennik, M., Kittel, P., Kloss, M., Twardy, J., Żurek, S., and Pawlyta, J.: Multiproxy study of anthropogenic and climatic changes in the last two millennia from a small mire in central Poland, in: Palaeolimnological Proxies as Tools of Environmental Reconstruction in Fresh Water, edited by: Buczkó, K., Korponai, J., Padisák, J., and Starratt, S. W., Springer Netherlands, Dordrecht, 213–230, https://doi.org/10.1007/978-90-481-3387-1_13, 2009a.
Lamentowicz, M., Milecka, K., Gałka, M., Cedro, A., Pawlyta, J., Piotrowska, N., Lamentowicz, Ł., and van der Knaap, W. O.: Climate and human induced hydrological change since AD 800 in an ombrotrophic mire in Pomerania (N Poland) tracked by testate amoebae, macro-fossils, pollen and tree rings of pine, Boreas, 38, 214–229, https://doi.org/10.1111/j.1502-3885.2008.00047.x, 2009b.
Lamentowicz, M., Lamentowicz, Ł., van der Knaap, W. O., Gąbka, M., and Mitchell, E. A. D.: Contrasting Species—Environment Relationships in Communities of Testate Amoebae, Bryophytes and Vascular Plants Along the Fen–Bog Gradient, Microb. Ecol., 59, 499–510, https://doi.org/10.1007/s00248-009-9617-6, 2010.
Lamentowicz, M., Gałka, M., Lamentowicz, Ł., Obremska, M., Kühl, N., Lücke, A., and Jassey, V. E. J.: Reconstructing climate change and ombrotrophic bog development during the last 4000 years in northern Poland using biotic proxies, stable isotopes and trait-based approach, Palaeogeography, Palaeoclimatology, Palaeoecology, 418, 261–277, https://doi.org/10.1016/j.palaeo.2014.11.015, 2015.
Lamentowicz, M., Gałka, M., Marcisz, K., Słowiński, M., Kajukało-Drygalska, K., Dayras, M. D., and Jassey, V. E. J.: Unveiling tipping points in long-term ecological records from Sphagnum-dominated peatlands, Biol. Lett., 15, 20190043, https://doi.org/10.1098/rsbl.2019.0043, 2019b.
Lamentowicz, M., Kołaczek, P., Mauquoy, D., Kittel, P., Łokas, E., Słowiński, M., Jassey, V. E. J., Niedziółka, K., Kajukało-Drygalska, K., and Marcisz, K.: Always on the tipping point – A search for signals of past societies and related peatland ecosystem critical transitions during the last 6500 years in N Poland, Quaternary Science Reviews, 225, 105954, https://doi.org/10.1016/j.quascirev.2019.105954, 2019a.
Lamentowicz, M., Andrews, L., Czerwiński, S., and Marcisz, K.: Multi-proxy palaeoecological studies from peatlands: a comprehensive review of recent advances and future developments, Earth-Science Reviews, 271, 105278, https://doi.org/10.1016/j.earscirev.2025.105278, 2025.
Limpens, J., Tomassen, H. B. M., and Berendse, F.: Expansion of Sphagnum fallax in bogs: striking the balance between N and P availability, Journal of Bryology, 25, 83–90, https://doi.org/10.1179/03736680235001733, 2003.
Loisel, J., Yu, Z., Beilman, D. W., Kaiser, K., and Parnikoza, I.: Peatland Ecosystem Processes in the Maritime Antarctic During Warm Climates, Sci. Rep., 7, 12344, https://doi.org/10.1038/s41598-017-12479-0, 2017.
Łuców, D., Küttim, M., Słowiński, M., Kołaczek, P., Karpińska-Kołaczek, M., Küttim, L., Salme, M., and Lamentowicz, M.: Searching for an ecological baseline: Long-term ecology of a post-extraction restored bog in Northern Estonia, Quaternary International, 607, 65–78, https://doi.org/10.1016/j.quaint.2021.08.017, 2022.
Lumbreras, A., Navarro, G., Pardo, C., and Molina, J. A.: Aquatic Ranunculus communities in the northern hemisphere: A global review, Plant Biosystems – An International Journal Dealing with all Aspects of Plant Biology, 145, 118–122, https://doi.org/10.1080/11263504.2011.602728, 2011.
Madeja, J. and Latowski, D.: Too Old AMS Radiocarbon Dates Obtained from Moss Remains from Lake Kwiecko Bottom Sediments (N Poland), Geochronometria, 32, 13–19, https://doi.org/10.2478/v10003-008-0029-2, 2008.
Magny, M., Bégeot, C., Guiot, J., and Peyron, O.: Contrasting patterns of hydrological changes in Europe in response to Holocene climate cooling phases, Quaternary Science Reviews, 22, 1589–1596, https://doi.org/10.1016/S0277-3791(03)00131-8, 2003.
Magny, M., Vannière, B., De Beaulieu, J.-L., Bégeot, C., Heiri, O., Millet, L., Peyron, O., and Walter-Simonnet, A.-V.: Early-Holocene climatic oscillations recorded by lake-level fluctuations in west-central Europe and in central Italy, Quaternary Science Reviews, 26, 1951–1964, https://doi.org/10.1016/j.quascirev.2006.04.013, 2007.
Marcisz, K., Fournier, B., Gilbert, D., Lamentowicz, M., and Mitchell, E. A. D.: Response of Sphagnum Peatland Testate Amoebae to a 1-Year Transplantation Experiment Along an Artificial Hydrological Gradient, Microb. Ecol., 67, 810–818, https://doi.org/10.1007/s00248-014-0367-8, 2014a.
Marcisz, K., Lamentowicz, Ł., Słowińska, S., Słowiński, M., Muszak, W., and Lamentowicz, M.: Seasonal changes in Sphagnum peatland testate amoeba communities along a hydrological gradient, European Journal of Protistology, 50, 445–455, https://doi.org/10.1016/j.ejop.2014.07.001, 2014b.
Marcisz, K., Tinner, W., Colombaroli, D., Kołaczek, P., Słowiński, M., Fiałkiewicz-Kozieł, B., Łokas, E., and Lamentowicz, M.: Long-term hydrological dynamics and fire history over the last 2000 years in CE Europe reconstructed from a high-resolution peat archive, Quaternary Science Reviews, 112, 138–152, https://doi.org/10.1016/j.quascirev.2015.01.019, 2015.
Marcisz, K., Jassey, V. E. J., Kosakyan, A., Krashevska, V., Lahr, D. J. G., Lara, E., Lamentowicz, Ł., Lamentowicz, M., Macumber, A., Mazei, Y., Mitchell, E. A. D., Nasser, N. A., Patterson, R. T., Roe, H. M., Singer, D., Tsyganov, A. N., and Fournier, B.: Testate Amoeba Functional Traits and Their Use in Paleoecology, Front. Ecol. Evol., 8, 575966, https://doi.org/10.3389/fevo.2020.575966, 2020a.
Marcisz, K., Kołaczek, P., Gałka, M., Diaconu, A.-C., and Lamentowicz, M.: Exceptional hydrological stability of a Sphagnum-dominated peatland over the late Holocene, Quaternary Science Reviews, 231, 106180, https://doi.org/10.1016/j.quascirev.2020.106180, 2020b.
Margielewski, W., Krąpiec, M., Buczek, K., Szychowska-Krąpiec, E., Korzeń, K., Niska, M., Stachowicz-Rybka, R., Wojtal, A. Z., Mroczkowska, A., Obidowicz, A., Sala, D., Drzewicki, W., Barniak, J., and Urban, J.: Hydrological variability of middle European peatland during the Holocene, inferred from subfossil bog pine and bog oak dendrochronology and high-resolution peat multiproxy analysis of the Budwity peatland (northern Poland), Science of The Total Environment, 931, 172925, https://doi.org/10.1016/j.scitotenv.2024.172925, 2024.
Mauquoy, D. and van Geel, B.: Mire and Peat Macros, in: Plant Macrofossil Methods and Studies, Elsevier, 2315–2336, https://doi.org/10.1016/B0-444-52747-8/00229-5, 2007.
Mauquoy, D. and Yeloff, D.: Raised peat bog development and possible responses to environmental changes during the mid- to late-Holocene. Can the palaeoecological record be used to predict the nature and response of raised peat bogs to future climate change?, Biodivers. Conserv., 17, 2139–2151, https://doi.org/10.1007/s10531-007-9222-2, 2008.
Mauquoy, D., van Geel, B., Blaauw, M., and van der Plicht, J.: Evidence from northwest European bogs shows “Little Ice Age” climatic changes driven by variations in solar activity, The Holocene, 12, 1–6, https://doi.org/10.1191/0959683602hl514rr, 2002.
Mauquoy, D., Yeloff, D., van Geel, B., Charman, D. J., and Blundell, A.: Two decadally resolved records from north-west European peat bogs show rapid climate changes associated with solar variability during the mid–late Holocene, J. Quaternary Science, 23, 745–763, https://doi.org/10.1002/jqs.1158, 2008.
Mauquoy, D., Hughes, P. D. M., and van Geel, B.: A protocol for plant macrofossil analysis of peat deposits, Mires and Peat, 7, 1–5, 2010.
Mayewski, P. A., Rohling, E. E., Curt Stager, J., Karlén, W., Maasch, K. A., Meeker, L. D., Meyerson, E. A., Gasse, F., van Kreveld, S., Holmgren, K., Lee-Thorp, J., Rosqvist, G., Rack, F., Staubwasser, M., Schneider, R. R., and Steig, E. J.: Holocene climate variability, Quat. Res., 62, 243–255, https://doi.org/10.1016/j.yqres.2004.07.001, 2004.
Mazei, Y. and Tsyganov, A. N.: Freshwater testate amoebae, KMK, Moscow, ISBN 5-87317-336-2, 2006.
McKay, N. P., Kaufman, D. S., Arcusa, S. H., Kolus, H. R., Edge, D. C., Erb, M. P., Hancock, C. L., Routson, C. C., Żarczyński, M., Marshall, L. P., Roberts, G. K., and Telles, F.: The 4.2 ka event is not remarkable in the context of Holocene climate variability, Nat. Commun., 15, 6555, https://doi.org/10.1038/s41467-024-50886-w, 2024.
Meisterfeld, R.: Testate Amoebae with Filopodia, The illustrated Guide to the Protozoa, 1054–1083, ISBN 1-891276-23-9, 2000.
Meisterfeld, R.: Testate amoebae, in: Patrimoines Naturels, edited by: Costello, M. J., Emblow, C. S., and White, R., Muséum National d'Histoire Naturelle – Institut d'Ecologie et de Gestion de la Biodiversité (I. E. G. B.) – Service du Patrimoine Naturel (S. P. N.), Paris, 54–57, ISBN 978-2-85653-538-7, 2001.
Montagnes, R. J. S.: The Habitat and Distribution of Meesia triquetra in North America and Greenland, The Bryologist, 93, 349, https://doi.org/10.2307/3243526, 1990.
Moore, P. A., Didemus, B. D., Furukawa, A. K., and Waddington, J. M.: Peat depth as a control on Sphagnum moisture stress during seasonal drought, Hydrological Processes, 35, e14117, https://doi.org/10.1002/hyp.14117, 2021.
Neukom, R., Steiger, N., Gómez-Navarro, J. J., Wang, J., and Werner, J. P.: No evidence for globally coherent warm and cold periods over the preindustrial Common Era, Nature, 571, 550–554, https://doi.org/10.1038/s41586-019-1401-2, 2019.
Noryśkiewicz, A.: Preliminary results of study on vegetation history in the Linje mire region using pollen analysis, Monographiae Botanicae, 94, 117–134, 2005.
Nungesser, M. K.: Modelling microtopography in boreal peatlands: hummocks and hollows, Ecological Modelling, 165, 175–207, https://doi.org/10.1016/S0304-3800(03)00067-X, 2003.
Ogden, C. G. and Hedley, R. H.: An Atlas of Freshwater Testate Amoebae, Oxford University Press, London, ISBN 0198585020, 1980.
Økland, K. A. and Økland, J.: Freshwater bryozoans (Bryozoa) of Norway: Distribution and ecology of Cristatella mucedo and Paludicella articulata, Hydrobiologia, 421, 1–24, https://doi.org/10.1023/A:1003917200848, 2000.
Oksanen, J., Simpson, G. L., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B., Solymos, P., Stevens, M. H. H., Szoecs, E., Wagner, H., Barbour, M., Bedward, M., Bolker, B., Borcard, D., Carvalho, G., Chirico, M., De Caceres, M., Durand, S., Evangelista, H. B. A., FitzJohn, R., Friendly, M., Furneaux, B., Hannigan, G., Hill, M. O., Lahti, L., McGlinn, D., Ouellette, M.-H., Ribeiro Cunha, E., Smith, T., Stier, A., Ter Braak, C. J. F., and Weedon, J.: vegan: Community Ecology Package, Comprehensive R Archive Network (CRAN), R Foundation for Statistical Computing, Vienna, Austria [code], https://doi.org/10.32614/CRAN.package.vegan, 2001.
Opravilová, V. and Hájek, M.: The Variation of Testacean Assemblages (Rhizopoda) Along the Complete Base-Richness Gradient in Fens: A Case Study from the Western Carpathians, Acta Protozool, 45, 191–204, 2006.
Pancost, R. D.: Biomarker carbon and hydrogen isotopes reveal changing peatland vegetation, hydroclimate and biogeochemical tipping points, Quaternary Science Reviews, 339, 108828, https://doi.org/10.1016/j.quascirev.2024.108828, 2024.
Payne, R. J. and Mitchell, E. A. D.: How many is enough? Determining optimal count totals for ecological and palaeoecological studies of testate amoebae, Journal of Paleolimnology, 42, 483–495, doi10.1007/s10933-008-9299-y, 2009.
Pazdur, A., Fontugne, M. R., and Pazdur, M. F.: Lateglacial and holocene water-level changes of the Gościa̧ż Lake, Central Poland, derived from carbon isotope studies of laminated sediment, Quaternary Science Reviews, 14, 125–135, https://doi.org/10.1016/0277-3791(94)00124-T, 1995.
Pędziszewska, A., Tylmann, W., Witak, M., Piotrowska, N., Maciejewska, E., and Latałowa, M.: Holocene environmental changes reflected by pollen, diatoms, and geochemistry of annually laminated sediments of Lake Suminko in the Kashubian Lake District (N Poland), Review of Palaeobotany and Palynology, 216, 55–75, https://doi.org/10.1016/j.revpalbo.2015.01.008, 2015.
Peters, M. E. and Higuera, P. E.: Quantifying the source area of macroscopic charcoal with a particle dispersal model, Quat. Res., 67, 304–310, https://doi.org/10.1016/j.yqres.2006.10.004, 2007.
Piilo, S. R., Väliranta, M. M., Amesbury, M. J., Aquino-López, M. A., Charman, D. J., Gallego-Sala, A., Garneau, M., Koroleva, N., Kärppä, M., Laine, A. M., Sannel, A. B. K., Tuittila, E., and Zhang, H.: Consistent centennial-scale change in European sub-Arctic peatland vegetation toward Sphagnum dominance—Implications for carbon sink capacity, Global Change Biology, 29, 1530–1544, https://doi.org/10.1111/gcb.16554, 2023.
Piilo, S. R., Kuoppamaa, M., Tahvanainen, T., Kumpula, T., Kuosmanen, N., Macias-Fauria, M., and Väliranta, M.: Drastic peatland regime shift and landscape disturbances connected to warm and cold climate events over the past centuries in subarctic Finland, Boreas, bor.70017, https://doi.org/10.1111/bor.70017, 2025.
Pisaric, M. F. J.: Long-distance transport of terrestrial plant material by convection resulting from forest fires, Journal of Paleolimnology, 28, 349–354, https://doi.org/10.1023/A:1021630017078, 2002.
Poolma, E., Marcisz, K., and Fiutek, P.: Plant macrofossil and testate-amoebae analysis for Linje peatland, Mendeley Data [data set], https://doi.org/10.17632/5f6565fdht.1, 2025.
Preislerová, Z., Marcenò, C., Loidi, J., Bonari, G., Borovyk, D., Gavilán, R. G., Golub, V., Terzi, M., Theurillat, J., Argagnon, O., Bioret, F., Biurrun, I., Campos, J. A., Capelo, J., Čarni, A., Çoban, S., Csiky, J., Ćuk, M., Ćušterevska, R., Dengler, J., Didukh, Y., Dítě, D., Fanelli, G., Fernández-González, F., Guarino, R., Hájek, O., Iakushenko, D., Iemelianova, S., Jansen, F., Jašková, A., Jiroušek, M., Kalníková, V., Kavgacı, A., Kuzemko, A., Landucci, F., Lososová, Z., Milanović, Đ., Molina, J. A., Monteiro-Henriques, T., Mucina, L., Novák, P., Nowak, A., Pätsch, R., Perrin, G., Peterka, T., Rašomavičius, V., Reczyńska, K., Rūsiòa, S., Mata, D. S., Guerra, A. S., Šibík, J., Škvorc, Ž., Stešević, D., Stupar, V., Świerkosz, K., Tzonev, R., Vassilev, K., Vynokurov, D., Willner, W., and Chytrý, M.: Structural, ecological and biogeographical attributes of European vegetation alliances, Applied Vegetation Science, 27, e12766, https://doi.org/10.1111/avsc.12766, 2024.
R Core Team: R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, https://www.R-project.org/ (last access: 27 October 2025), 2024.
Ralska-Jasiewiczowa, M. (Ed.): Lake Gościąż, Central Poland: A Monographic Study, W. Szafer Institute of Botany, Polish Academy of Sciences, ISBN 83-85444-64-5, 1998.
Reczuga, M. K., Lamentowicz, M., Mulot, M., Mitchell, E. A. D., Buttler, A., Chojnicki, B., Słowiński, M., Binet, P., Chiapusio, G., Gilbert, D., Słowińska, S., and Jassey, V. E. J.: Predator–prey mass ratio drives microbial activity under dry conditions in Sphagnum peatlands, Ecology and Evolution, 8, 5752–5764, https://doi.org/10.1002/ece3.4114, 2018.
Reimer, P. J., Austin, W. E. N., Bard, E., Bayliss, A., Blackwell, P. G., Bronk Ramsey, C., Butzin, M., Cheng, H., Edwards, R. L., Friedrich, M., Grootes, P. M., Guilderson, T. P., Hajdas, I., Heaton, T. J., Hogg, A. G., Hughen, K. A., Kromer, B., Manning, S. W., Muscheler, R., Palmer, J. G., Pearson, C., Van Der Plicht, J., Reimer, R. W., Richards, D. A., Scott, E. M., Southon, J. R., Turney, C. S. M., Wacker, L., Adolphi, F., Büntgen, U., Capano, M., Fahrni, S. M., Fogtmann-Schulz, A., Friedrich, R., Köhler, P., Kudsk, S., Miyake, F., Olsen, J., Reinig, F., Sakamoto, M., Sookdeo, A., and Talamo, S.: The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP), Radiocarbon, 62, 725–757, https://doi.org/10.1017/RDC.2020.41, 2020.
Roland, T. P., Caseldine, C. J., Charman, D. J., Turney, C. S. M., and Amesbury, M. J.: Was there a “4.2 ka event” in Great Britain and Ireland? Evidence from the peatland record, Quaternary Science Reviews, 83, 11–27, https://doi.org/10.1016/j.quascirev.2013.10.024, 2014.
Ronkainen, T., McClymont, E. L., Tuittila, E.-S., and Väliranta, M.: Plant macrofossil and biomarker evidence of fen–bog transition and associated changes in vegetation in two Finnish peatlands, The Holocene, 24, 828–841, https://doi.org/10.1177/0959683614530442, 2014.
Rundgren, M., Kokfelt, U., Schoning, K., and Wastegård, S.: Holocene wet shifts in NW European bogs: evidence for the roles of external forcing and internal feedback from a high-resolution study of peat properties, plant macrofossils and testate amoebae, J. Quaternary Science, 38, 423–439, https://doi.org/10.1002/jqs.3485, 2023.
Ruppel, M., Väliranta, M., Virtanen, T., and Korhola, A.: Postglacial spatiotemporal peatland initiation and lateral expansion dynamics in North America and northern Europe, The Holocene, 23, 1596–1606, https://doi.org/10.1177/0959683613499053, 2013.
Rydin, H., Gunnarsson, U., and Sundberg, S.: The Role of Sphagnum in Peatland Development and Persistence, in: Boreal Peatland Ecosystems, vol. 188, edited by: Wieder, R. K. and Vitt, D. H., Springer, Berlin, Heidelberg, 47–65, https://doi.org/10.1007/978-3-540-31913-9_4, 2006.
Samson, M., Słowińska, S., Słowiński, M., Lamentowicz, M., Barabach, J., Harenda, K., Zielińska, M., Robroek, B. J. M., Jassey, V. E. J., Buttler, A., and Chojnicki, B. H.: The Impact of Experimental Temperature and Water Level Manipulation on Carbon Dioxide Release in a Poor Fen in Northern Poland, Wetlands, 38, 551–563, https://doi.org/10.1007/s13157-018-0999-4, 2018.
Seppä, H. and Birks, H. J. B.: July mean temperature and annual precipitation trends during the Holocene in the Fennoscandian tree-line area: pollen-based climate reconstructions, The Holocene, 11, 527–539, https://doi.org/10.1191/095968301680223486, 2001.
Seppä, H., Birks, H. J. B., Giesecke, T., Hammarlund, D., Alenius, T., Antonsson, K., Bjune, A. E., Heikkilä, M., MacDonald, G. M., Ojala, A. E. K., Telford, R. J., and Veski, S.: Spatial structure of the 8200 cal yr BP event in northern Europe, Clim. Past, 3, 225–236, https://doi.org/10.5194/cp-3-225-2007, 2007.
Seppä, H., Bjune, A. E., Telford, R. J., Birks, H. J. B., and Veski, S.: Last nine-thousand years of temperature variability in Northern Europe, Clim. Past, 5, 523–535, https://doi.org/10.5194/cp-5-523-2009, 2009.
Siemensma, F. J.: Microworld, world of amoeboid organisms, World-wide electronic publication, Kortenhoef, the Netherlands, https://www.arcella.nl (last access: 27 October 2025), 2024.
Sillasoo, U., Mauquoy, D., Blundell, A., Charman, D., Blaauw, M., Daniell, J. R. G., Toms, P., Newberry, J., Chambers, F. M., and Karofeld, E.: Peat multi-proxy data from Männikjärve bog as indicators of late Holocene climate changes in Estonia, Boreas, 36, 20–37, https://doi.org/10.1111/j.1502-3885.2007.tb01177.x, 2007.
Sim, T. G., Swindles, G. T., Morris, P. J., Baird, A. J., Charman, D. J., Amesbury, M. J., Beilman, D., Channon, A., and Gallego-Sala, A. V.: Ecology of peatland testate amoebae in Svalbard and the development of transfer functions for reconstructing past water-table depth and pH, Ecological Indicators, 131, 108122, https://doi.org/10.1016/j.ecolind.2021.108122, 2021a.
Sim, T. G., Swindles, G. T., Morris, P. J., Baird, A. J., Cooper, C. L., Gallego-Sala, A. V., Charman, D. J., Roland, T. P., Borken, W., Mullan, D. J., Aquino-López, M. A., and Gałka, M.: Divergent responses of permafrost peatlands to recent climate change, Environ. Res. Lett., 16, 034001, https://doi.org/10.1088/1748-9326/abe00b, 2021b.
Sim, T. G., Swindles, G. T., Morris, P. J., Baird, A. J., Gallego-Sala, A. V., Wang, Y., Blaauw, M., Camill, P., Garneau, M., Hardiman, M., Loisel, J., Väliranta, M., Anderson, L., Apolinarska, K., Augustijns, F., Aunina, L., Beaulne, J., Bobek, P., Borken, W., Broothaerts, N., Cui, Q.-Y., Davies, M. A., Ejarque, A., Farrell, M., Feeser, I., Feurdean, A., Fewster, R. E., Finkelstein, S. A., Gaillard, M.-J., Gałka, M., Heffernan, L., Hoevers, R., Jones, M., Juselius-Rajamäki, T., Karofeld, E., Knorr, K.-H., Korhola, A., Kupriyanov, D., Kylander, M. E., Lacourse, T., Lamentowicz, M., Lavoie, M., Lemdahl, G., Łuców, D., Magnan, G., Maksims, A., Mansilla, C. A., Marcisz, K., Marinova, E., Mathijssen, P. J. H., Mauquoy, D., Mazei, Y. A., Mazei, N., McCarroll, J., McCulloch, R. D., Milner, A. M., Miras, Y., Mitchell, F. J. G., Novenko, E., Pelletier, N., Peros, M. C., Piilo, S. R., Pilote, L.-M., Primeau, G., Rius, D., Robin, V., Robitaille, M., Roland, T. P., Ryberg, E., Sannel, A. B. K., Schittek, K., Servera-Vives, G., Shotyk, W., Słowiński, M., Stivrins, N., Swinnen, W., Thompson, G., Tiunov, A., Tsyganov, A. N., Tuittila, E.-S., Verstraeten, G., Wallenius, T., Webb, J., Willard, D., Yu, Z., Zaccone, C., and Zhang, H.: Regional variability in peatland burning at mid-to high-latitudes during the Holocene, Quaternary Science Reviews, 305, 108020, https://doi.org/10.1016/j.quascirev.2023.108020, 2023.
Słowińska, S., Słowiński, M., and Lamentowicz, M.: Relationships between Local Climate and Hydrology in Sphagnum Mire: Implications for Palaeohydrological Studies and Ecosystem Management, Polish Journal of Environmental Studies, 4, 779–787, 2010.
Słowińska, S., Słowiński, M., Marcisz, K., and Lamentowicz, M.: Long-term microclimate study of a peatland in Central Europe to understand microrefugia, Int. J. Biometeorol., 66, 817–832, https://doi.org/10.1007/s00484-022-02240-2, 2022.
Słowiński, M.: Macrofossil reconstruction of preboreal wetland formed on dead ice block: a case study of the Borzechowo mire in East Pomerania, Poland, Studia Quaternaria, 27, 3–10, 2010.
Słowiński, M., Błaszkiewicz, M., Brauer, A., Noryśkiewicz, B., Ott, F., and Tyszkowski, S.: The role of melting dead ice on landscape transformation in the early Holocene in Tuchola Pinewoods, North Poland, Quaternary International, 388, 64–75, https://doi.org/10.1016/j.quaint.2014.06.018, 2015.
Słowiński, M., Marcisz, K., Płóciennik, M., Obremska, M., Pawłowski, D., Okupny, D., Słowińska, S., Borówka, R., Kittel, P., Forysiak, J., Michczyńska, D. J., and Lamentowicz, M.: Drought as a stress driver of ecological changes in peatland – A palaeoecological study of peatland development between 3500 BCE and 200 BCE in central Poland, Palaeogeography, Palaeoclimatology, Palaeoecology, 461, 272–291, https://doi.org/10.1016/j.palaeo.2016.08.038, 2016.
Słowiński, M., Zawiska, I., Ott, F., Noryśkiewicz, A. M., Plessen, B., Apolinarska, K., Rzodkiewicz, M., Michczyńska, D. J., Wulf, S., Skubała, P., Kordowski, J., Błaszkiewicz, M., and Brauer, A.: Differential proxy responses to late Allerød and early Younger Dryas climatic change recorded in varved sediments of the Trzechowskie palaeolake in Northern Poland, Quaternary Science Reviews, 158, 94–106, https://doi.org/10.1016/j.quascirev.2017.01.005, 2017.
Stastney, P. and Black, S.: Bog Microtopography and the Climatic Sensitivity of Testate Amoeba Communities: Implications for Transfer Function-Based Paleo-Water Table Reconstructions, Microb. Ecol., 80, 309–321, https://doi.org/10.1007/s00248-020-01499-5, 2020.
Stivrins, N.: Climate change impact on peatland dynamics during the Holocene in Latvia, Northeastern Europe, Catena, 254, https://doi.org/10.1016/j.catena.2025.108965, 2025.
Succow, M. and Joonsten, H. (Eds.): Landschaftsökologische Moorkunde, Schweizerbart Science Publishers, Stuttgart, Germany, ISBN 978-3-510-65198-6, 2012.
Swindles, G. T., Blundell, A., Roe, H. M., and Hall, V. A.: A 4500 year proxy climate record from peatlands in the North of Ireland: the identification of widespread summer “drought phases”?, Quaternary Science Reviews, 29, 1577–1589, https://doi.org/10.1016/j.quascirev.2009.01.003, 2010.
Swindles, G. T., Morris, P. J., Baird, A. J., Blaauw, M., and Plunkett, G.: Ecohydrological feedbacks confound peat-based climate reconstructions, Geophysical Research Letters, 39, 2012GL051500, https://doi.org/10.1029/2012GL051500, 2012.
Swindles, G. T., Morris, P. J., Mullan, D. J., Payne, R. J., Roland, T. P., Amesbury, M. J., Lamentowicz, M., Turner, T. E., Gallego-Sala, A., Sim, T., Barr, I. D., Blaauw, M., Blundell, A., Chambers, F. M., Charman, D. J., Feurdean, A., Galloway, J. M., Gałka, M., Green, S. M., Kajukało, K., Karofeld, E., Korhola, A., Lamentowicz, Ł., Langdon, P., Marcisz, K., Mauquoy, D., Mazei, Y. A., McKeown, M. M., Mitchell, E. A. D., Novenko, E., Plunkett, G., Roe, H. M., Schoning, K., Sillasoo, Ü., Tsyganov, A. N., van der Linden, M., Väliranta, M., and Warner, B.: Widespread drying of European peatlands in recent centuries, Nat. Geosci., 12, 922–928, https://doi.org/10.1038/s41561-019-0462-z, 2019.
Swindles, G. T., Roland, T. P., Amesbury, M. J., Lamentowicz, M., McKeown, M. M., Sim, T. G., Fewster, R. E., and Mitchell, E. A. D.: Quantifying the effect of testate amoeba decomposition on peat-based water-table reconstructions, European Journal of Protistology, 74, 125693, https://doi.org/10.1016/j.ejop.2020.125693, 2020.
Swindles, G. T., Mullan, D. J., Brannigan, N. T., Fewster, R. E., Sim, T. G., Gallego-Sala, A., Blaauw, M., Lamentowicz, M., Jassey, V. E. J., Marcisz, K., Green, S. M., Roland, T. P., Loisel, J., Amesbury, M. J., Blundell, A., Chambers, F. M., Charman, D. J., Evans, C. R. C., Feurdean, A., Galloway, J. M., Gałka, M., Karofeld, E., Keaveney, E. M., Korhola, A., Lamentowicz, Ł., Langdon, P., Mauquoy, D., McKeown, M. M., Mitchell, E. A. D., Plunkett, G., Roe, H. M., Turner, T. E., Sillasoo, Ü., Väliranta, M., van der Linden, M., and Warner, B.: Climate and water-table levels regulate peat accumulation rates across Europe, PLoS One, 20, e0327422, https://doi.org/10.1371/journal.pone.0327422, 2025.
Szambelan, W., Niebieszczański, J., Karpińska-Kołaczek, M., Lamentowicz, M., Marcisz, K., Leszczyńska, K., Poolma, E., Amon, L., Veski, S., and Kołaczek, P.: Tracing the environmental footprint of a Lusatian Urnfield culture stronghold in northern Poland, Journal of Archaeological Science: Reports, 67, 105408, https://doi.org/10.1016/j.jasrep.2025.105408, 2025.
Tallis, J. H. and Birks, H. J. B.: The Past and Present Distribution of Scheuchzeria Palustris L. in Europe, The Journal of Ecology, 53, 287, https://doi.org/10.2307/2257976, 1965.
Theurer, T., Mauquoy, D., Hadden, R., Muirhead, D., Campbell-Lochrie, Z., Córdoba, S. V., Von Scheffer, C., and Coathup, D. T.: A novel proxy for energy flux in multi-era wildfire reconstruction, Sci. Rep., 14, 26409, https://doi.org/10.1038/s41598-024-78219-3, 2024.
Timmermann, T.: Sphagnum-Moore in Nordostbrandenburg: stratigraphisch-hydrodynamische Typisierung und Vegetationswandel seit 1923, PhD thesis, Technischen Univesität Berlin, ISBN 978-3-443-64217-4, 1998.
Timmermann, T.: Hydrologische Dynamik von Kesselmooren und ihre Bedeutung für die Gehölzentwicklung, Telma, 33, 85–107, 2003.
Timmermann, T.: Wasserstandsschwankungen und Vegetationsdynamik in Kesselmooren: interne und externe Faktoren., in: Aktuelle Probleme im Wasserhaushalt von Nordostdeutschland: Trends, Ursachen, Lösungen, edited by: Kaiser, K., Libra, J., Merz, B., Bens, O., and Hüttl, R. F., Scientific Technical Report 10/10, Deutsches GeoForschungsZentrum, 218–223, https://doi.org/10.2312/GFZ.b103-10106, 2010.
Timmermann, T. and Succow, M.: Kesselmoore, Landschaftsökologische Moorkunde, 379–390, ISBN 9783510654543, 2001.
Tinner, W., Hofstetter, S., Zeugin, F., Conedera, M., Wohlgemuth, T., Zimmermann, L., and Zweifel, R.: Long-distance transport of macroscopic charcoal by an intensive crown fire in the Swiss Alps – implications for fire history reconstruction, The Holocene, 16, 287–292, https://doi.org/10.1191/0959683606hl925rr, 2006.
Tomlinson, P.: An aid to the identification of fossil buds, bud-scales and catkin-scales of British trees and shrubs, Circaea. Association for Environmental Archaeology, 3, 45–130, 1985.
Turetsky, M. R., Weston, D. J., Cox, W. D., Petro, C., and Shaw, A. J.: The challenging but unique eco-evolutionary aspects of Sphagnum moss, New Phytologist, 247, 1608–1621, https://doi.org/10.1111/nph.70233, 2025.
Turner, T. E., Swindles, G. T., and Roucoux, K. H.: Late Holocene ecohydrological and carbon dynamics of a UK raised bog: impact of human activity and climate change, Quaternary Science Reviews, 84, 65–85, https://doi.org/10.1016/j.quascirev.2013.10.030, 2014.
United Nations Environment Programme: Global Peatlands Assessment: The State of the World's Peatlands – Evidence for Action toward the Conservation, Restoration, and Sustainable Management of Peatlands, United Nations Environment Programme, https://doi.org/10.59117/20.500.11822/41222, 2022.
Väliranta, M., Korhola, A., Seppä, H., Tuittila, E.-S., Sarmaja-Korjonen, K., Laine, J., and Alm, J.: High-resolution reconstruction of wetness dynamics in a southern boreal raised bog, Finland, during the late Holocene: a quantitative approach, The Holocene, 17, 1093–1107, https://doi.org/10.1177/0959683607082550, 2007.
Väliranta, M., Blundell, A., Charman, D. J., Karofeld, E., Korhola, A., Sillasoo, Ü., and Tuittila, E.-S.: Reconstructing peatland water tables using transfer functions for plant macrofossils and testate amoebae: A methodological comparison, Quaternary International, 268, 34–43, https://doi.org/10.1016/j.quaint.2011.05.024, 2012.
Väliranta, M., Salonen, J. S., Heikkilä, M., Amon, L., Helmens, K., Klimaschewski, A., Kuhry, P., Kultti, S., Poska, A., Shala, S., Veski, S., and Birks, H. H.: Plant macrofossil evidence for an early onset of the Holocene summer thermal maximum in northernmost Europe, Nat. Commun., 6, 6809, https://doi.org/10.1038/ncomms7809, 2015.
van Breemen, N.: How Sphagnum bogs down other plants, TREE, 10, 270–275, https://doi.org/10.1016/0169-5347(95)90007-1, 1995.
van der Linden, M. and van Geel, B.: Late Holocene climate change and human impact recorded in a south Swedish ombrotrophic peat bog, Palaeogeography, Palaeoclimatology, Palaeoecology, 240, 649–667, https://doi.org/10.1016/j.palaeo.2006.03.039, 2006.
Verhoeven, J. T. A.: Wetlands in Europe: Perspectives for restoration of a lost paradise, Ecological Engineering, 66, 6–9, https://doi.org/10.1016/j.ecoleng.2013.03.006, 2014.
Veski, S., Seppä, H., and Ojala, A. E. K.: Cold event at 8200 yr B. P. recorded in annually laminated lake sediments in eastern Europe, Geol., 32, 681, https://doi.org/10.1130/G20683.1, 2004.
Wanner, H., Solomina, O., Grosjean, M., Ritz, S. P., and Jetel, M.: Structure and origin of Holocene cold events, Quaternary Science Reviews, 30, 3109–3123, https://doi.org/10.1016/j.quascirev.2011.07.010, 2011.
Wanner, H., Mercolli, L., Grosjean, M., and Ritz, S. P.: Holocene climate variability and change; a data-based review, JGS, 172, 254–263, https://doi.org/10.1144/jgs2013-101, 2015.
Yan, M. and Liu, J.: Physical processes of cooling and mega-drought during the 4.2 ka BP event: results from TraCE-21ka simulations, Clim. Past, 15, 265–277, https://doi.org/10.5194/cp-15-265-2019, 2019.
Zander, P. D., Żarczyński, M., Tylmann, W., Vogel, H., and Grosjean, M.: Subdecadal Holocene Warm-Season Temperature Variability in Central Europe Recorded by Biochemical Varves, Geophysical Research Letters, 51, e2024GL110871, https://doi.org/10.1029/2024GL110871, 2024a.
Żurek, S.: Abiotic natural environment in the area of selected raised mires, Monographiae Botanicae, 94, 19–36, 2005.