the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 27 Oct 2025
| 27 Oct 2025
Quantitative reconstruction of deglacial bottom-water nitrate in marginal Pacific seas using the pore density of denitrifying benthic foraminifera
Anjaly Govindankutty Menon
Aaron L. Bieler
Hanna Firrincieli
Rachel Alcorn
Niko Lahajnar
Catherine V. Davis
Ralf Schiebel
Dirk Nürnberg
Gerhard Schmiedl
Nicolaas Glock
Quantifying past ocean nitrate concentrations is crucial for understanding the global nitrogen cycle. Here, we reconstruct deglacial bottom-water nitrate concentrations ([NO]BW) in the oxygen-deficient zones of the Sea of Okhotsk, the Gulf of California, the Mexican Margin, and the Gulf of Guayaquil. Using the pore density of denitrifying benthic foraminifera as a nitrate proxy, differences in [NO]BW are observed at the study sites spanning the Last Glacial Maximum to the Holocene. Changes in water-column denitrification, water-mass ventilation, primary productivity, and sea surface temperatures may account for nitrate differences at the study sites. The [NO]BW in the Sea of Okhotsk, the Gulf of California, and the Gulf of Guayaquil are influenced by the intermediate water masses while, the [NO]BW at the Mexican Margin is likely influenced by deglacial changes in the Pacific Deep Water. The comparison of past and present [NO] shows that the modern Gulf of Guayaquil and the Gulf of California currently have stronger oxygen-deficient zones with higher denitrification than during the Last Glacial Maximum. In contrast, the modern Mexican Margin and the Sea of Okhotsk may have higher oxygen than during the Last Glacial Maximum, indicated by low modern denitrification.
- Article
(2272 KB) - Full-text XML
-
Supplement
(649 KB) - BibTeX
- EndNote




The marine nitrogen cycle is a complex web of microbially mediated processes controlling the inventory and distribution of bioavailable nitrogen in marine environments (Casciotti, 2016). Biological nitrogen fixation by nitrogen-fixing diazotrophs (e.g., cyanobacteria) in the surface layer is the main source of bioavailable nitrogen in the ocean, and denitrification and anammox, are the main fixed nitrogen loss processes (Lam and Kuypers, 2011), both of which occur under low-oxygen conditions. The primary form of bioavailable nitrogen in the ocean is nitrate (NO), (Casciotti, 2016), which is a limiting nutrient throughout the tropical and subtropical oceans (Moore et al., 2013).
Oxygen-deficient zones (ODZs) are regions of very low dissolved oxygen (O2) where the O2 concentration is less than 22 µmol kg−1, usually within depths of 100–1200 m (Levin, 2003; 2018). Oxygen plays a key role in the marine nitrogen cycle (Keeling et al., 2010) because some microbial processes require oxygen while others are inhibited by it (Voss et al., 2013). For example, denitrification (reduction of nitrate to dinitrogen gas) in the ocean occurs in suboxic (oxygen <5 µmol kg−1) conditions (Codispoti et al., 2001; Levin, 2018). On a global scale, ∼ 30 %–50 % of fixed nitrogen loss in the world's oceans occurs in ODZs (Gruber, 2008), either through denitrification or anammox (Devol et al., 2006; Lam and Kuypers, 2011; Evans et al., 2023). Due to the complex interactions and feedbacks within the biogeochemical nitrogen cycle, the amount of benthic denitrification also influences other important processes, such as global nitrogen fixation and net primary production (Somes et al., 2017; Li et al., 2024). Oxygen Deficient Zones cover only 1 % of the world's seafloor (Codispoti et al., 2001), however, 10 % of the global benthic denitrification occurs in these regions (Bohlen et al., 2012). Observations and climate model simulations have predicted that ODZs will continue to expand until at least the year 2100 (Stramma et al., 2008, 2010; Schmidtko et al., 2017; Oschlies, 2021). However, the long-term evolution of ODZs remains uncertain (Yamamoto et al., 2015; Takano et al., 2018; Fu et al., 2018; Frölicher et al., 2020). There is growing evidence that ODZs may contract during transient and equilibrium climate warmings over timescales of millennia and beyond (Auderset et al., 2022; Moretti et al., 2024). Considering the role of ODZs in modulating the marine nitrogen cycle, it is of key scientific interest to understand how nitrogen cycling works in these ecosystems and the potential factors that influence the nitrogen cycle.
In this study, we use the pore density (number of pores per unit area) of Bolivina spissa and Bolivina subadvena as a NO proxy (Fig. 1a) to reconstruct bottom-water nitrate [NO]BW in intermediate water depths of the Sea of Okhotsk, the Gulf of California, the Gulf of Guayaquil, and in the Pacific Deep Water (PDW) depths of the Mexican Margin (Figs. 2 and 3). The [NO]BW calibration using the pore density of B. spissa and B. subadvena (see Fig. 1b) developed in Govindankutty Menon et al. (2023) is applied in the current study. Combining a proxy for [NO]BW (pore density of denitrifying foraminifera) and a proxy for N-cycle processes in the water column (δ15Nbulk) facilitates a more comprehensive understanding of past N-cycling in different zones of the water column. Here, we try to understand 1) whether there are differences in reconstructed [NO]BW between today, deglacial, and glacial periods in the four studied sites, and 2) whether the reconstructed [NO]BW records are in agreement with insights drawn from δ15Nbulk data.
1.1 Application of δ15Nbulk and its potential limitations
The stable isotope signature of nitrogen in the sedimentary organic matter (δ15Nbulk) is an established proxy for water-column denitrification and for understanding changes associated with nutrient utilization (Thunell et al., 2004; Robinson et al., 2009; Martinez and Robinson, 2010; Dubois et al., 2011, 2014; Tesdal et al., 2013; Wang et al., 2019; Riechelson et al., 2024). An increase (or decrease) in nutrient availability in relation to nutrient demand results in an increase (or decrease) in δ15N values (Wada and Hattori, 1978; Montoya et al., 1990). When the oxygen in the ocean is depleted, either due to global warming or increased remineralization, denitrification rates in the water column are also increasing and so is δ15N (Wang et al., 2019). Therefore, δ15Nbulk can be an important tool for reconstructing past changes in denitrification in the ODZs.
The δ15N records from the bulk sediment can be subject to interlinked processes/or sources which can complicate their interpretation. For example, diagenetic alteration during sinking in the water column and burial in the sediment (Altabet and Francois, 1994; Lourey et al., 2003), as well as terrestrial or shelf sources of organic and inorganic nitrogen (Schubert and Calvert, 2001; Kienast et al., 2005; Meckler et al., 2011), and remotely advected water masses with different δ15N values (for e.g., Southern Californian margin; Liu and Kaplan, 1989), could influence the δ15N signatures in sediments. Nevertheless, Tesdal et al. (2013) proposed that δ15Nbulk can be a reliable indicator for individual locations reflecting the oceanographic conditions of the surrounding environments.
The nitrogen isotopes of organic matter bound and protected within the calcite shell of planktic foraminifera (δ15NFB) are less subjected to diagenesis or sedimentary contamination than δ15Nbulk and can be used to understand major nitrogen transformations occurring in the ocean (Ren et al., 2012; Studer et al., 2021). There are well-documented disagreements between bulk sediment δ15N and foraminifera-bound δ15N records, particularly in glacial-interglacial comparisons (Studer et al., 2021). While δ15Nbulk suggests strong variability in water-column denitrification between the LGM and deglaciation, δ15NFB records indicate a more moderate change, with a peak during deglaciation but relatively stable values during the LGM and Holocene. This highlights that δ15Nbulk and δ15NFB may reflect different aspects of the nitrogen cycle (Studer et al., 2021). Recent studies (Auderset et al., 2022; Hess et al., 2023; Moretti et al., 2024) based on δ15NFB have shown that water column denitrification decreased and ODZs contracted during warmer-than-present periods of the Cenozoic. In contrast, Riechelson et al. (2024) used δ15Nbulk and hypothesized that the decrease in δ15Nbulk values over the Holocene is related to a decrease in Southern Ocean nutrient utilization and not due to a decrease in denitrification.
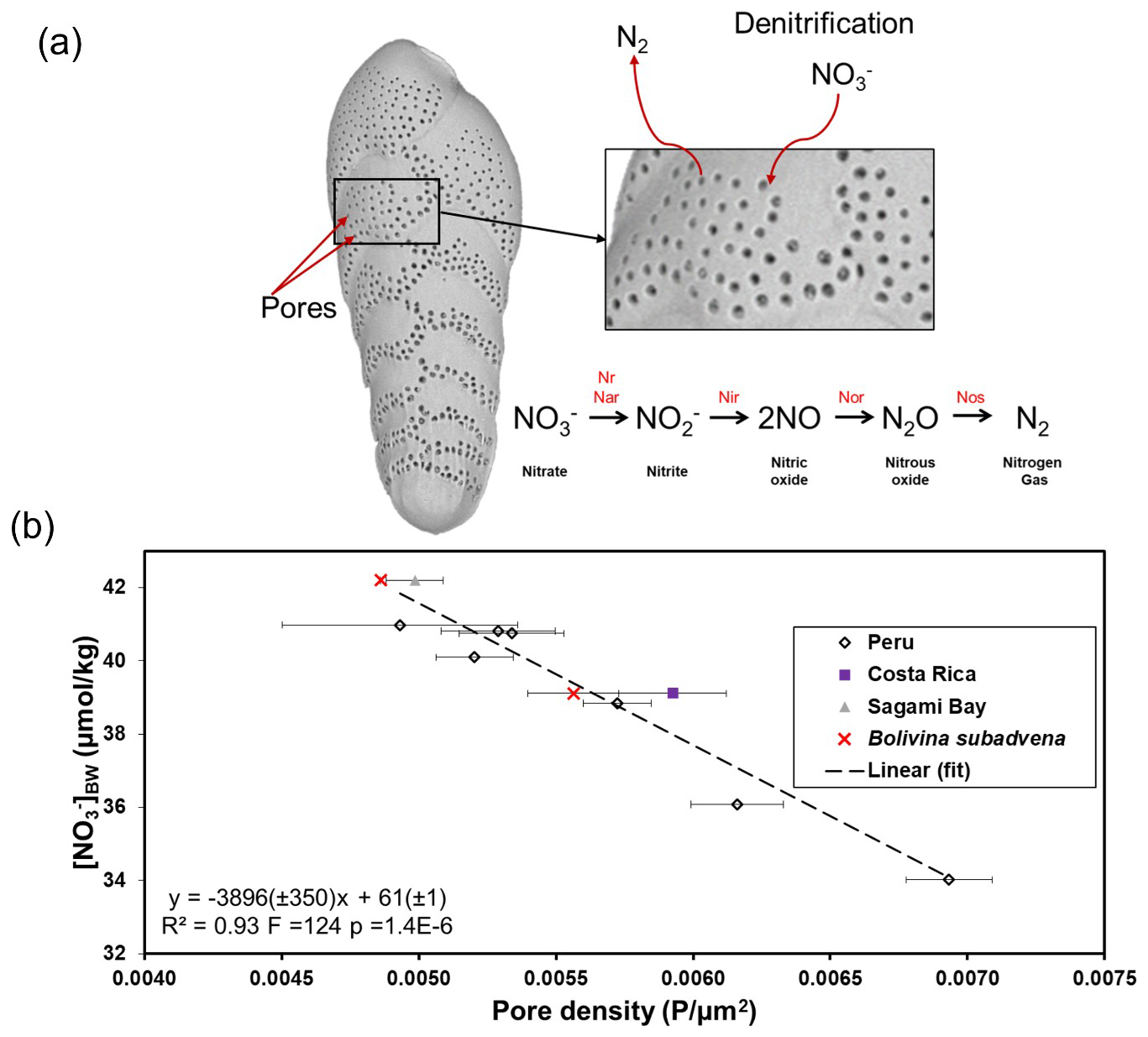
Figure 1The (a) schematic view of nitrate (NO) uptake, and the excretion of nitrogen gas (N2) by the benthic foraminifera Bolivina spissa. The step-wise denitrification pathway from NO to N2 involving enzymes such as nitrate reductase (Nr Nar), nitrite reductase (Nir), nitric-oxide reductase (Nor), and nitrous oxide reductase (Nos) is also shown. (b) Correlation between pore density of Bolivina spissa from Peru, off Costa Rica, Sagami Bay, and Bolivina subadvena with bottom-water nitrate [NO]BW from Govindankutty Menon et al. (2023). If no species name is indicated in the legend, the analysed species was B. spissa. The error bars are 1 standard error of the mean.
1.2 Pore density of benthic foraminifera as a bottom-water nitrate proxy
Foraminifera account for a major part of benthic denitrification in the ODZs (up to 100 % in some environments) (Piña-Ochoa et al., 2010a, b; Glock et al., 2013; Dale et al., 2016; Choquel et al., 2021; Rakshit et al., 2025). Some species, for example B. spissa, which are abundant in ODZs in and around the Pacific Ocean (Glock et al., 2011; Fontanier et al., 2014) can use NO as an electron acceptor (see Fig. 1a) and thus can denitrify (Risgaard-Petersen et al., 2006; Piña-Ochoa et al., 2010a, b). A study by Glock et al. (2019) proposed for some denitrifying foraminifera, denitrification is their preferred respiration pathway. The uptake of NO by these foraminifera is likely through pores in the test (see Fig. 1a). Nitrate is completely denitrified to dinitrogen gas (N2) partly by the foraminifera themselves (Risgaard-Petersen et al., 2006; Woehle and Roy et al., 2018; Orsi et al., 2020; Gomaa et al., 2021), and partly supported by prokaryotic endobionts (Bernhard et al., 2012a, Woehle and Roy et al., 2022). To date, benthic foraminifera are the only eukaryote holobiont known to perform complete heterotrophic denitrification (Risgaard-Petersen et al., 2006; Kamp et al., 2015). Every Bolivina species tested so far (including Bolivina seminuda), can denitrify (Piña-Ochoa et al., 2010a; Bernhard et al., 2012b), suggesting that denitrification is a common survival strategy of Bolivinidae under oxygen-depleted conditions (Glock et al., 2019). This makes species of this genus particularly suitable candidates for reconstructing past nitrate levels using pore characteristics as a proxy. In low-oxygen environments, such as the ODZs off Peru, Costa Rica, and the hypoxic Sagami Bay, B. spissa increase their pore density with decreasing ambient NO availability (Govindankutty Menon et al., 2023). Thus, the pore density of several Bolivina species, such as B. spissa, and B. subadvena, is an empirically calibrated proxy that shows the strongest correlation with the bottom-water nitrate concentration (see Fig. 1b) (Glock et al., 2011; Govindankutty Menon et al., 2023) rather than bottom-water oxygen, temperature, water depth, salinity or pore water nitrate.
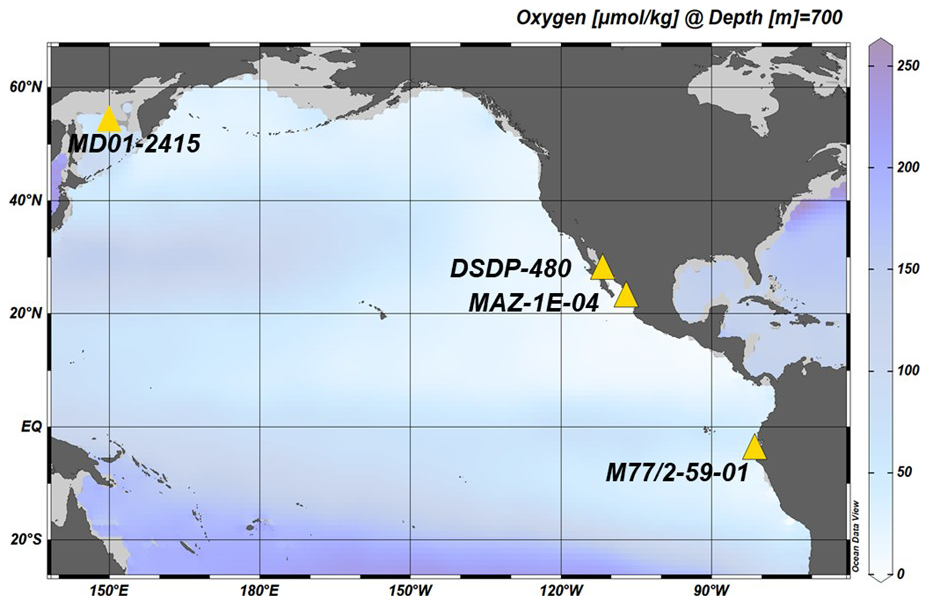
Figure 2Location of sediment cores used in the current study and mean annual oxygen concentrations at 700 m depth (Garcia et al., 2019). Sediment cores are indicated by yellow triangles: Sea of Okhotsk (core MD01-2415; water depth: 822 m), Gulf of California (DSDP Site- 480; water depth: 747 m), Mexican Margin (core MAZ-1E-04; water depth: 1463 m), and Gulf of Guayaquil (core M77/2-59-01; water depth: 997 m). Map created with Ocean Data View (Schlitzer, 2023).
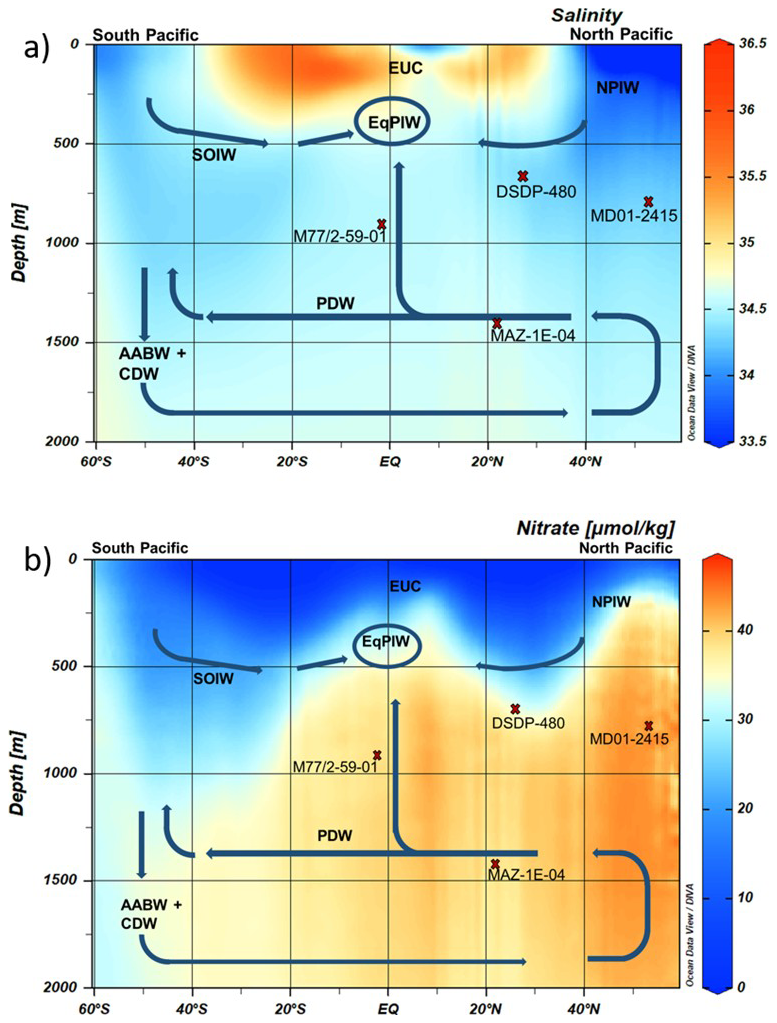
Figure 3Modern (a) salinity and (b) nitrate distribution along a N–S transect across the Pacific (Garcia et al., 2019) with major subsurface and deep-water masses (blue arrows) and formation areas of North Pacific Intermediate Water (NPIW) and Southern Ocean Intermediate Water (SOIW) are included. Sediment cores used for [NO]BW reconstruction are shown (red crosses) projected to the N-S hydrographic transect. Equatorial Pacific Intermediate Water (EqPIW), Equatorial Undercurrent (EUC), NPIW, SOIW, Pacific Deep Water (PDW), Antarctic Bottom Water (AABW), and Circumpolar Deep Water (CDW). Profiles generated by Ocean Data View (Schlitzer, 2023) using the data from World Ocean Atlas 2018 (Garcia et al., 2019).
2.1 Study area and sampling of sediment cores
We used downcore samples from the Eastern Tropical South Pacific, ETSP (Gulf of Guayaquil (M77/2-59-01), Eastern Tropical North Pacific the ETNP (Mexican Margin, MAZ-1E-04), the Gulf of California (Guaymas Basin, DSDP-64-480), and the Sea of Okhotsk (MD01-2415), over the last ∼ 20 000 years (Fig. 3). The Gulf of Guayaquil sediment core M77/2-59-01 (03°57.01′ S, 81°19.23′ W, recovery 13.59 m) was collected from the northern edge of the ODZ at a water depth of 997 m during the RV Meteor cruise M77/2 in 2008 (Mollier-Vogel et al., 2013, 2019; Nürnberg et al., 2015). The piston core MAZ-1E-04, Mexican Margin (22.9° N, 106.91° W) was collected on board the RV El Puma at a water depth of 1463 m. The CALYPSO giant piston core MD01-2415 (53°57.09′ N, 149°57.52′ E, recovery 46.23 m) was recovered from the northern slope of the Sea of Okhotsk at 822 m water depth during the WEPAMA cruise MD122 of the RV Marion Dufresne (Holbourn et al., 2002; Nürnberg and Tiedemann, 2004). The Deep-Sea Drilling Project core DSDP-480 (27°54′ N, 111°39′ W) from the Gulf of California was retrieved at a water depth of 747 m close to the Guaymas Basin. For details on the sampling procedure of foraminiferal specimens, please refer to the Supplement Methods section in the Supplement.
2.2 Automated image analysis
All specimens of B. spissa and B. subadvena were imaged using a Scanning Electron Microscope (Hitachi Tabletop SEM TM4000 series) at Hamburg University, Germany with an accelerating voltage of 15 kV using a back-scattered electron (BSE) detector (Further methodological details are provided in the Supplement).
Following the image analysis, pore density data of benthic foraminifera from the four ODZs were used for the quantitative reconstruction of [NO]BW (Fig. 4). We distinguished five different time intervals, including the Last Glacial Maximum (LGM; 22–17 ka BP), Heinrich Stadial 1 (H1; 17–15 ka BP), Bølling – Allerød (BA; 14.7–12.9 ka BP), Younger Dryas (YD; 12.9–11.7 ka BP), Early Holocene (EH; 11.7–8.2 ka BP) and Middle to Late Holocene (MLH; 8–0 ka BP) to describe the [NO]BW in the East Pacific and the Sea of Okhotsk. We present updated chronostratigraphies of the studied cores, primarily based on accelerator mass spectrometry (AMS) radiocarbon (14C) dating, as detailed in the Supplement Methods.
The [NO]BW from all cores were calculated using the calibration equation;
where PD is the pore density of benthic foraminifera (Govindankutty Menon et al., 2023).
The standard error of the mean (SEM) for one sample was calculated using the equation;
where n is the number of specimens analyzed in each sample and SD is 1 standard deviation of mean reconstructed [NO]BW.
A complete error propagation was done for the calculation of the errors of the reconstructed [NO3]BW including both the uncertainty of the mean PD within the samples and the uncertainties of the calibration function. The reconstructed [NO]BW and the calculated SEM and SD of each sample are shown in the Supplement.
2.3 Sedimentary nitrogen isotope (δ15Nbulk) measurements
We have measured sedimentary nitrogen isotope (δ15Nbulk) rather than δ15NFB from cores taken from the Sea of Okhotsk, and Gulf of California, because the low abundances of foraminifera were utilized for other analysis. The analysis of bulk sediments allows for high-resolution records. Prior to the δ15Nbulk measurements, the Total Nitrogen (TN %) content of 20 sediment samples from the Sea of Okhotsk and 54 samples from the Gulf of California were measured at the Institute for Geology, Hamburg University, Germany using a flash combustion method with a Eurovector EA-3000 analyzer. The δ15Nbulk measurements for both the Sea of Okhotsk and the Gulf of California were accomplished at the Max Planck Institute for Chemistry (Mainz), Germany using a DELTA V ADVANTAGE Isotope Ratio Mass Spectrometer (IRMS) equipped with a FLASH 2000 Organic Elemental Analyzer. The results were expressed in standard δ-notation (Eq. 4). The standard deviation (± SD) of all individual analysis runs based on a certified international reference standard (USGS65) and internal laboratory standards (L-Phenylalanine and L-Glutamic acid) referenced to certified international reference standards was <0.3 ‰. The δ15Nbulk data for the Sea of Okhotsk and the Gulf of California are shown in Supplement Table S1.
For the Gulf of Guayaquil core M77/2-59-01, the δ15Nbulk data published by Mollier-Vogel et al. (2019) was used. Their measurements were done on ∼ 5–50 mg of homogenized and freeze-dried bulk sediments using a Carlo-Erba CN analyzer 2500 interfaced directly to a Micromass-Isoprime mass spectrometer at Bordeaux University. Results are expressed in standard δ-notation (Eq. 4) relative to atmospheric dinitrogen gas (N2).
2.4 Nitrate offset to present conditions
The reconstructed [NO]BW from each location is subtracted from the modern [NO] present at the respective locations from similar water depths the cores were retrieved from. This provided the [NO] offset which is the difference (Δ[NO] (µM)) between the modern [NO] and the past reconstructed [NO]BW. The modern [NO] for each location was taken from World Ocean Atlas 2018 (Garcia et al., 2019). The details are given in the Supplement.
We reconstructed deglacial [NO]BW using downcore sediment samples from the Sea of Okhotsk (MD01-2415), the Gulf of California (DSDP- 480), the Mexican Margin (MAZ-1E-04), and the Gulf of Guayaquil (M77/2-59-01). The reconstructed [NO]BW was compared to δ15Nbulk records of all cores (Fig. 4). All data records presented cover the time period starting from the Last Glacial Maximum, except for the core from the Sea of Okhotsk, which covers the late deglacial to the Holocene.
3.1 Sea of Okhotsk (MD01-2415)
The Sea of Okhotsk core MD01-2415 covers the Younger Dryas, (YD, 12.8 ka BP) until the Middle to Late Holocene (MLH, 4.9 ka BP). The reconstructed [NO]BW values range from 32.8 to 44.1 µmol kg−1 (Fig. 4a). A gradual increase in [NO]BW is observed from the Younger Dryas to the Middle to Late Holocene. At the beginning of the Younger Dryas at 12.8 ka BP, [NO]BW were relatively high and then decreased to a minimum value of 32.8 µmol kg−1 at 12.4 ka BP. Since then, [NO]BW steadily increased until the Middle to Late Holocene (MLH, 44.1 µmol kg−1) (Fig. 4a). The [NO]BW during the Middle to Late Holocene (mean =41.2 µmol kg−1) is significantly (t-test, p=0.023) higher than during the Younger Dryas (mean =36.7 µmol kg−1). The sedimentary δ15Nbulk record covers the interval from the Late Heinrich Stadial 1 (H1, 15.4 ka BP) to the Middle Holocene (6.1 ka BP). The δ15Nbulk values were relatively high ranging from 7.1 ‰ to 9.4 ‰ with an average of 8.7 ‰. The δ15Nbulk values increased steadily from the Late Heinrich Stadial 1 (15.4 ka BP) to the Early Holocene (EH, 10 ka BP) with higher values centered between the Late Younger Dryas (11.9 ka BP) and the beginning of the Early Holocene. Since then, the δ15Nbulk values decreased until the Middle to Late Holocene.
3.2 Gulf of California (DSDP-480)
The analyzed sections of DSDP Site 480 covered the Last Glacial Maximum (22 ka BP) until the Early Holocene (10.8 ka BP). The reconstructed [NO]BW ranged from 41.4 to 49.1 µmol kg−1. The highest [NO]BW of 49.1 µmol kg−1 occurred during the Last Glacial Maximum (18.2 ka BP). The data points from the Early Holocene (11.6–10.8 ka BP) were the only Holocene data from this core providing the lowest [NO]BW estimate of 42.1 µmol kg−1 during the Early Holocene (10.8 ka BP) (Fig. 4b). A distinct difference in [NO]BW between the glacial period (mean =46.1 µmol kg−1) and the Early Holocene (42.7 µmol kg−1) was observed with [NO]BW found to be substantially higher during the glacial period (t-test, p=0.0067) (Fig. 4b). Accordingly, the [NO]BW followed a decreasing pattern from the glacial period to the Early Holocene. The δ15Nbulk values varied between 6.4 ‰ and 13 ‰ with an average of 10.2 ‰ (Fig. 4b). The δ15Nbulk values from the Guaymas Basin were similar to the δ15Nbulk values (average 9.6 ‰) of Pride (1997) and Altabet et al. (1999). During the last glacial period, the δ15Nbulk values were low ranging from 8.5 ‰ to 9 ‰. At the onset of the deglaciation, the δ15Nbulk values increased by more than 2 ‰ with large-scale changes reaching a maximum of 13 ‰ during the Younger Dryas. Afterward, we observed a gradual decline in δ15Nbulk values throughout the Middle to Late Holocene (mean 10.7 ‰) and this pattern continued to the present.
3.3 Mexican Margin (MAZ-1E-04)
This core MAZ-1E-04 covered the Last Glacial Maximum (20.5 ka BP) until the Early Holocene (10.47 ka BP). The [NO]BW values range from 37.7 to 43.5 µmol kg−1. We observed the highest [NO]BW during the Younger Dryas. From the beginning to the end of the Last Glacial Maximum, [NO]BW followed a decreasing trend (Fig. 4c). The [NO]BW levels continued to steadily decrease until Heinrich Stadial 1 and consistently stayed low throughout this period. There was a strong change in [NO]BW from the end of Heinrich Stadial 1 to the end of Younger Dryas (Fig. 4c). We observed a peak in [NO]BW from the beginning of Bølling-Allerød, BA (14.29 ka BP) and it continued throughout the Younger Dryas (Fig. 4c). Afterwards, [NO]BW declined during the Early Holocene. The δ15Nbulk values taken from Alcorn et al. (2025) followed an increasing trend from the glacial towards the deglacial period (Fig. 4c).
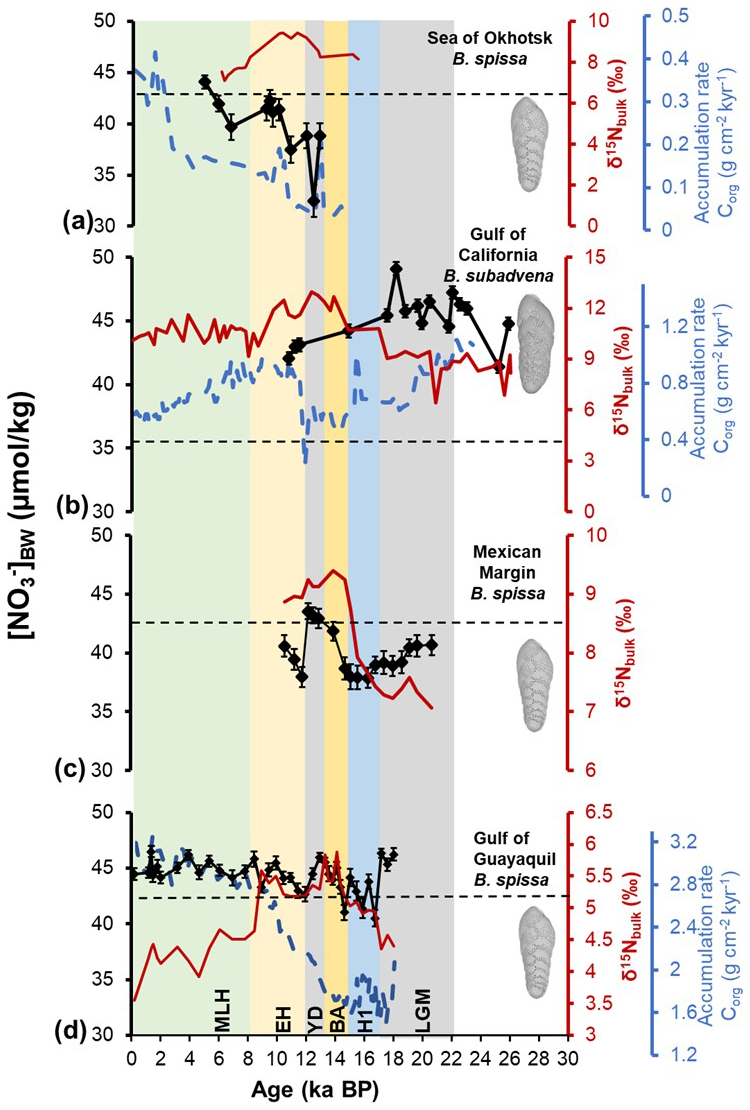
Figure 4Quantitative [NO]BW reconstruction using the pore density of fossil specimens of B. spissa, B. subadvena from (a) the Sea of Okhotsk (MD01-2415), (b) the Gulf of California (DSDP-480), (c) the Mexican Margin (MAZ-1E-04), and (d) Gulf of Guayaquil (M77/2-59-01). The sedimentary nitrogen isotope (δ15Nbulk) records from the Sea of Okhotsk, and the Gulf of California are measured in this study, and the Gulf of Guayaquil is from Mollier-Vogel et al. (2019), and the Mexican Margin δ15Nbulk data is from Alcorn et al. (2025). The error bars of [NO]BW represent 1 SEM including a complete error propagation (using Eqs. 3 and 4). The accumulation rate of total organic carbon (Supplement) calculated from published literature (Bubenshchikova et al., 2015; Leclaire and Kerry, 1982; Mollier-Vogel et al., 2019) is shown in blue dashed lines for the Sea of Okhotsk, the Gulf of California and the Gulf of Guayaquil cores respectively. The black dashed lines indicate the modern nitrate concentration of each location. Time intervals Middle to Late Holocene (MLH), Early Holocene (EH), Younger Dryas (YD), Bølling-Allerød (BA), Heinrich Stadial 1 (H1), and Last Glacial Maximum (LGM) are shown in the figure.
3.4 Gulf of Guayaquil (M77/2-59-01)
This core covered the Last Glacial Maximum (18 ka BP) until the Middle to Late Holocene (0.18 ka BP). The reconstructed [NO]BW values range from 40.5 to 46.5 µmol kg−1. The highest [NO]BW occurred during the Last Glacial Maximum (Fig. 4d). The reconstructed [NO]BW levels during the Last Glacial Maximum (mean =45.6 µmol kg−1) were slightly higher than during the Middle to Late Holocene (mean =44.9 µmol kg−1) (t-test, p=0.046). The δ15Nbulk values were relatively low ranging between 4 ‰ and 6 ‰ (Fig. 4d). During the Last Glacial Maximum, the δ15Nbulk values were low, varying between 4.4 ‰ and 4.6 ‰, close to the typical mean range of dissolved nitrate in the ocean (Sigman et al., 1997). Subsequently, the δ15Nbulk values increased from 16.7 ka BP (4.9 ‰), where we observed a decline in [NO]BW to 8.9 ka BP (5.6 ‰). The highest δ15Nbulk values centered at ∼ 14 ka BP (5.9 ‰). From 8.9 ka BP onwards, a long-term decrease in δ15Nbulk (<4.4 ‰) was observed until the Latest Holocene, consistent with higher [NO]BW levels during the Holocene (Fig. 4d). Despite higher [NO]BW levels, our reconstruction doesn't show any strong variations during the Holocene.
The pore density of benthic foraminifera represents a promising but still developing proxy for reconstructing past nitrate dynamics. Like most proxies based on biology, it reflects an indirect physiological response rather than a direct measure of nitrate. In addition, species-specific variability requires careful taxonomic control or its interpretation carries inherent limitations especially since not many records are available, yet, for this proxy. Thus, we used a multiproxy approach and combined it with δ15Nbulk, which provides a complementary perspective that strengthens reconstructions of nitrogen-cycling processes in oxygen-deficient zones.
4.1 Sea of Okhotsk
Our data show that [NO]BW levels gradually increased through time and reached modern concentrations during the Middle-Holocene (Fig. 4a). Most of the nutrients in the northwestern Pacific including the Sea of Okhotsk are supplied by the upwelling of the North Pacific Deep Water (NPDW) (Gorbarenko et al., 2014). The weakened Kuroshio current (Ujiié and Ujiié, 1999) and increased sea ice extent (Ternois et al., 2001) weakened the upwelling of NPDW during the Last Glacial Maximum (LGM). Subsequent studies (Gray et al., 2020; Rae et al., 2020) have shown that the expansion of the North Pacific Gyre also resulted in less upwelling of NPDW during the LGM.
During the LGM, the subpolar North Pacific was better ventilated at intermediate depths (Keigwin, 1998) and export productivity was reduced (Ternois et al., 2001; Narita et al., 2002; Seki et al., 2004). This is consistent with a strengthened meridional overturning circulation, with enhanced formation of intermediate waters and advection of nutrient-depleted subtropical waters to high latitudes (Rae et al., 2020). Furthermore, the North Pacific subpolar gyre extended ∼ 3° further south during the LGM (Gray et al., 2020), which shifted the westerly winds southward. This may have resulted in less upwelling of the NPDW during the LGM.
The prolonged ice cover with low biological productivity (Ternois et al., 2001; Narita et al., 2002; Seki et al., 2004; Rae et al., 2020) and well-oxygenated water masses (Keigwin, 1998) might have prevented the formation of an oxygen deficient zone (ODZ) in the Sea of Okhotsk (Bubenshchikova et al., 2015). This is supported by the absence of B. spissa, which are adapted to living in dysoxic conditions, in our records during the LGM.
Deglacial low [NO]BW which correspond to higher δ15Nbulk values (Fig. 4a) could be due to enhanced primary productivity. It is important to note, however, that δ15Nbulk is influenced by diagenetic alteration and the incorporation of allochthonous nitrogen, which can obscure the local denitrification signal. Therefore, interpretations of δ15Nbulk trends should be made cautiously and ideally corroborated with complementary proxies, such as foraminiferal pore density. Increased nutrient supply from the Asian continental shelves and sea-ice retreat (Ternois et al., 2001) strengthened primary productivity. Indeed, the accumulation rate of total organic carbon was relatively higher during the Younger Dryas (Bubenshchikova et al., 2015) in our core (Fig. 4a). The increased oxygen demand and weakened ventilation of intermediate waters in the subarctic Pacific (Lembke-Jene et al., 2018) gradually intensified the ODZ. These poorly oxygenated conditions conceivably strengthened denitrification, resulting in low deglacial [NO]BW levels. However, during the Middle to Late Holocene (MLH) a reorganization in atmospheric circulation favored enhanced formation of oxygenated North Pacific Intermediate Water (NPIW) (Wang et al., 2020). Thus, mid-depth ventilation was closely associated with atmospheric circulation in the Holocene and a weakened ODZ (Ohkushi et al., 2013; Bubenshchikova et al., 2015; Wang et al., 2020). These rising oxygen concentrations probably reduced denitrification (low δ15Nbulk) in the Sea of Okhotsk, resulting in higher [NO]BW comparable to today's conditions (Fig. 4). The δ15Nbulk values show a maximum from 13 ka to 10 ka BP, which indicates increased water-column denitrification during that time. Nevertheless, the [NO]BW increased during this time, which indicates a decoupling from denitrification in the oxygen minimum in the water column and the [NO]BW. This could be related to the sea level rise during that time (Waelbroeck et al., 2008), which increased the vertical distance of the sediments (i.e., bottom water) at the sampling site from the center of denitrification.
4.2 Gulf of California
The Gulf of California ODZ is influenced by both intermediate and deep-water properties, similar to that of the open Pacific Ocean. Thus, the ODZ intensity in the Guaymas Basin is largely dependent on the oxygen content and ventilation of inflowing NPIW from the Sea of Okhotsk (Pride et al., 1999) and the demand for oxygen at depth. During the glacial period, the dissolved oxygen concentrations were higher due to better-ventilated NPIW at intermediate depths of the Northeast Pacific (Keigwin and Jones, 1990; Ganeshram et al., 1995; Keigwin 1998; Duplessy et al., 1988; Herguera et al., 2010; Cartapanis et al., 2011). Modeling studies show that the Laurentide and Cordilleran ice sheets increased in size (Benson et al., 2003), lowering the temperature of North America (Romanova et al., 2006) during the glacial period. The cold sinking air over the ice sheet established a semi-permanent high-pressure cell (Kutzbach and Wright Jr., 1985; Romanova et al., 2006) causing a substantially weaker North Pacific High (Ganeshram and Pedersen, 1998) or the southward displacement of the Inter Tropical Convergence Zone (Cheshire and Thurow, 2013). This resulted in a weak California Current along the coast and reduced upwelling-favorable winds (Cartapanis et al., 2011) along the North American coastline and reduced primary productivity (Ganeshram and Pedersen, 1998; Hendy et al., 2004; Cartapanis et al., 2011; Chang et al., 2015) within the ETNP and the Gulf of California during the glacial period. The nitrogen isotope ratios in the Guaymas Basin can be affected by subsurface denitrification in the Gulf and in the ETNP (Pride et al 1999). The increase in dissolved oxygen during the glacial period might have reduced water column denitrification (low δ15Nbulk) thereby increasing the [NO]BW (Fig. 4b).
Our study finds a declining trend in reconstructed [NO]BW during the Early Holocene, slowly approaching modern concentrations. This coincides with a maximum in δ15Nbulk values, suggesting elevated denitrification. This agrees with previous studies in the ETNP (Kienast et al., 2002) and within the Gulf of California (Pride et al., 1999), which showed that high denitrification most likely was associated with warming temperatures that occurred during this period. Furthermore, the scarcity of benthic foraminifera after the Early Holocene in our study coincides with laminations of the sediment core (Keigwin and Jones, 1990) below 10.8 ka BP, where reconstructed [NO]BW begins to decrease. It is possible that redox conditions were too hostile for benthic foraminifers in the time periods when laminated sediments formed. We acknowledge limitations in our Holocene reconstruction due to the low abundance of B. subadvena and the limited calibration dataset available for this species, which may introduce a systematic offset (Govindankutty Menon et al., 2023). Bolivina subadvena was used in this core due to the unavailability of B. spissa, and some values fall outside the existing calibration range. We also cannot rule out other factors influencing the proxy signal, such as microhabitat variability. Additional data and further proxy calibration are therefore essential to improve the robustness of Holocene bottom-water nitrate reconstructions.
4.3 Mexican Margin
Our study finds a steep rise in [NO]BW between the Bølling-Allerød (BA) and the Younger Dryas (YD) (Fig. 4c). The transition period from the BA to the Holocene involved rapid oxygenation with increased oxygen levels at the onset of the YD (Jaccard and Galbraith, 2012; Ohkushi et al., 2013; Taylor et al., 2017). This has been linked to active ventilation by increased NPIW production at high latitudes in the North Pacific (Van Geen et al., 1996; Emmer and Thunell, 2000; Okazaki et al., 2010; Cartapanis et al., 2011; Chang et al., 2014). In addition, there was low primary productivity (Hendy et al., 2004; Pospelova et al., 2015), and a higher influx of freshwater (Broecker et al., 1985; Clark et al., 2002) during the YD. However, considering the deep location of the Mexican Margin core below the direct influence of intermediate water masses (Fig. 3), it is less likely to be reflected in the [NO]BW. Bulk sediment δ15N records in the ETNP (Ganeshram et al., 1995; Pride et al., 1999; Emmer and Thunell, 2000; Kienast et al., 2002; Hendy et al., 2004) found a decrease in δ15Nbulk during the YD due to reduced denitrification. Furthermore, a foraminifera-bound nitrogen isotope (δ15NFB) study (Studer et al., 2021) in the eastern tropical Pacific also found a decrease in δ15NFB signatures during the Younger Dryas (Fig. 4c). In contrast to this, a bulk sediment δ15N record of MAZ-1E-04 (Alcorn et al., 2025) depicts an increase in water column denitrification during the Younger Dryas. Thus, reduced denitrification may not be the dominant factor that led to the elevated [NO]BW during this time. Instead, the Mexican Margin may be more influenced by the NO variability from the Pacific Deep Water, PDW (see Fig. 3). Deep-sea reorganization and ventilation during the deglaciation may have influenced the [NO]BW. At the onset of the deglaciation, deep Southern Ocean ventilation (reduced 14C ventilation ages) and atmospheric carbon dioxide (CO2) synchronously increased (Robinson et al., 2009; Burke and Robinson, 2012; Rae et al., 2018). This deglacial increase in 14C ventilation in the Pacific Ocean suggests that most of the increase in atmospheric CO2 is derived from old carbon in the Southern and Pacific Oceans (Rafter et al., 2022). The increase in reconstructed [NO]BW during the YD may thus reflect the release of sequestered nutrient- and carbon dioxide-rich waters during the deglaciation (Robinson et al., 2009; Rafter et al., 2022).
The relatively high [NO]BW during the glacial period (Fig. 4c), before its decline in Heinrich Stadial 1, is likely indicative of reduced water-column denitrification (Ganeshram et al., 1995, 2000) due to reduced productivity (Ganeshram et al., 1995; Ganeshram and Pedersen, 1998) and low organic matter flux through the oxygen minimum zone (Ganeshram et al., 2000). In the ETNP, including the Mexican Margin, coastal upwelling is driven by trade winds generated by subtropical high-pressure centers. These high-pressure centers largely result from differential heating of the land and the ocean. As a result of glacial cooling on land, these high-pressure systems and the associated trade winds that drive the upwelling have likely been weakened (Ganeshram and Pedersen, 1998).
4.4 Gulf of Guayaquil
The core M77/2-59-01 is in a region that is sensitive to changes in subsurface denitrification in the ETSP (Robinson et al., 2007, 2009; Dubois et al., 2011, 2014). The elevated reconstructed [NO]BW levels (Fig. 4d) during the glacial period suggest decreased water-column denitrification (Salvatteci et al., 2014; Erdem et al., 2020; Glock et al., 2022) and relatively low local productivity (Ganeshram et al., 2000; Robinson et al 2007, 2009; Martinez and Robinson et al., 2010; Salvatteci et al., 2016). Nutrient export to the deep Southern Ocean waters increased due to the sluggish Atlantic Meridional Overturning Circulation (Skinner et al., 2010), and increased atmospheric iron (Fe) deposition (Somes et al., 2017) during the glacial period. This reduced the transport of preformed NO to the tropics via the Subantarctic Mode Water (SAMW), limiting productivity. In fact, the total organic carbon (Fig. 4d) depicts low productivity during this period. Furthermore, the colder sea surface temperature (SST) and the accelerated formation of SAMW and Antarctic Intermediate Water masses (Russell and Dickson, 2003; Galbraith et al., 2004) and the stronger high-latitude winds in the Southern Hemisphere (Karstensen and Quadfasel, 2002) increased the ventilation rate (Meissner et al., 2005; Jaccard and Galbraith, 2012; Muratli et al., 2010) during the glacial period. The resulting increased oxygen concentrations (Robinson et al., 2005; Robinson et al., 2007) decreased the volume of ODZs, and nitrogen loss processes (lower δ15Nbulk values, Fig. 4d) during the glacial period. In addition, enhanced Fe deposition (Somes et al., 2017), and the glacial low sea level (Clark and Mix, 2002; Wallmann et al., 2016), may have influenced the nitrate inventory in the tropical and subtropical southern hemisphere.
A study by Glock et al. (2018) on core M77/2-52-2 from Peru applying the pore density of B. spissa also shows elevated [NO]BW during the Last Glacial Maximum, a similar decline in [NO]BW during the Heinrich Stadial 1 and thereafter a steady decrease in [NO]BW throughout the Holocene.
The deglacial decline in [NO]BW, especially during Heinrich Stadial 1 in this study (Fig. 4d), indicates a gradual increase in surface productivity and bottom-water deoxygenation. High export production strengthened the expansion of the ETSP ODZ during the deglaciation as compared to LGM and MLH (Salvatteci et al., 2016; Glock et al., 2018; Mollier-Vogel et al., 2019). This is consistent with the denitrification signal in the Eastern Equatorial Pacific through westward advection from the Southeast Pacific margins (Martinez and Robinson, 2010).
The shift towards generally higher reconstructed [NO]BW from the Middle-Holocene, (Fig. 4d), implies a profound change in the climatic state of the Peruvian upwelling system and the associated ODZ during this time. From the deglaciation toward the Late Holocene, there was a general increase in productivity (Mollier-Vogel et al., 2019) as shown by organic carbon accumulation rates (Fig. 4d). This increase in organic matter input and/or preservation was likely related to an increase in upwelling-driven delivery of nutrients towards the surface. The gradual decrease in δ15Nbulk values and higher [NO]BW was likely related to a relaxation in nutrient utilization with a nutrient supply exceeding the biological demand (Riechelson et al., 2024). Moreover, the core M77/2-59-01 was retrieved outside of the core ODZ and is under the strong influence of the oxygen and nutrient-rich Equatorial Under Current subsurface waters (Salvatteci et al., 2019; Mollier-Vogel et al., 2019). These waters might have ventilated the Northern Peruvian margin and deepened the oxycline at this site during the Middle-Holocene. Furthermore, enhanced zonal SST (Koutavas et al., 2006) and a northward shift of the ITCZ strengthened the Pacific Walker and Hadley circulation during the Middle-Holocene across the tropical Pacific (Koutavas et al., 2006; Mollier-Vogel et al., 2013; Salvatteci et al., 2019). These enhanced atmospheric circulations brought oxygen-rich waters to intermediate depths off Peru via the equatorial subsurface countercurrents (Koutavas et al., 2006; Mollier-Vogel et al., 2013; Salvatteci et al., 2019). Hence, increased ventilation of subsurface water masses reduced the strength of nitrogen loss processes and nutrient uptake during the MLH.
At present, the only quantitative reconstruction of bottom-water oxygen from these locations is the core M77/2-59-01 from the Gulf of Guayaquil reported by Erdem et al. (2020). Their record suggests a decline in bottom-water oxygen from the deglacial period to the Holocene. Future more detailed comparisons of the nitrate reconstructions with quantitative bottom water oxygen records at the same cores will further improve our understanding about variability in redox conditions and nitrogen cycling.
4.5 Comparison of past and present [NO] at the studied locations
The [NO] during the present and past are compared to assess the resilience of our chosen study locations towards environmental and ecological impacts of climate change. The generally positive Δ[NO] that we found (Fig. 4b) in the Gulf of California (Guaymas Basin) and the Gulf of Guayaquil indicate that today the [NO] is lower than in the past. This suggests that today the nitrogen loss processes at these two core sites are stronger, most likely related to ocean warming and a decline in oxygen concentration of bottom waters. The Gulf of California core is within the heart of the oxygen-deficient zone, and thus changes in ODZ oxygenation or denitrification will be more evident in this core than in any other core studied. Under nitrogen limitation, negative feedbacks (e.g., anammox) result in a decline in productivity (Naafs et al., 2019; Wallmann et al., 2022), which will stabilize the oxygen concentration. In the case of the Gulf of California, sediments are enriched in reactive iron (Fe) (Scholz et al., 2019). The decreasing NO concentrations in the bottom water reduce the flux of NO into the surface sediment. This leads to the release of sedimentary Fe, which enhances nitrogen fixation in the Guaymas Basin (Scholz et al., 2019). Thus, increased denitrification might not act as negative feedback in the Gulf of California because it might be countered by increased nitrogen fixation (White et al., 2013).
In the case of the Gulf of Guayaquil (Fig. 4d), whether today's elevated denitrification could enhance N2 fixation also depends on the availability of Fe (Pennington et al., 2006). The primary productivity of the Peruvian ODZ is Fe limited due to the reduction of particular Fe oxides in shelf and slope sediments (Scholz et al., 2014). Modeling studies show that primary productivity will be amplified in the Peruvian ODZ due to the release of Fe from shelf and slope sediments (Wallmann et al., 2022). This may induce deoxygenation and drive the expansion and intensification of Peruvian ODZ resulting in a positive feedback loop, like in the Gulf of California. This situation is indicated by lower [NO] today compared to the past ∼ 20 000 years.
The negative nitrate Δ[NO] in the Sea of Okhotsk and the Mexican Margin (Fig. 4a and c) indicates that modern [NO] levels are higher than in the reconstructed past. This suggests that modern nitrogen loss is decreased at these two core sites compared to the last deglaciation. The higher modern [NO] in the Sea of Okhotsk is likely associated with less primary productivity and more oxygen in the water column similar to the situation established in the MLH. The higher modern [NO]BW in the case of the Mexican Margin could be associated with sea level rise. The ODZ in the Mexican Continental Margin might have shifted to shallower depths today with less/or no benthic denitrification in intermediate water depths at the core site, resulting in high [NO]BW levels. During the glacial period, continental shelves were exposed due to sea-level lowstands (Clark and Mix, 2002; Kuhlmann et al., 2004; Wallmann et al., 2016), the main areas of primary productivity may have migrated offshore from the shallow shelf towards the continental slope relative to their Holocene positions. A similar situation occurred at the Benguela upwelling system during the LGM: TOC accumulation at the continental slope increased during the LGM in response to the seaward shift of centers of enhanced productivity (Mollenhauer et al., 2002). This offshore shift of the productivity centers and the most likely reduced remineralization rates, due to lower temperatures, indicate that the center of the ODZ at the Mexican Margin before sea level rise was possibly deeper than today. However, with the deglacial eustatic sea-level rise, the ODZ may have shifted to shallower depths. This shifted the main zone of denitrification further away from the seafloor, resulting in the increased modern [NO]BW in comparison to the LGM.
The quantitative reconstruction of [NO]BW using the pore density of denitrifying benthic foraminifera over the last deglaciation at the four studied ODZs provides a comprehensive understanding of the past [NO]. The Gulf of Guayaquil and Gulf of California data shows elevated [NO]BW during the glacial period compared to deglacial and modern conditions. Considering the well-ventilated intermediate water masses in the Sea of Okhotsk, the Sea of Okhotsk may have also elevated [NO]BW in the glacial period. For the Mexican Margin core, [NO]BW was particularly strong during the Younger Dryas. The reconstructed [NO]BW from the Sea of Okhotsk, the Gulf of California, and the Gulf of Guayaquil are influenced by the formation of the North Pacific Intermediate Water. However, the [NO]BW in the deeper site, the Mexican Margin is likely influenced by the NO variability in Pacific Deep Water. The modern Gulf of Guayaquil and the Gulf of California have low [NO] associated with increased denitrification and a strengthening ODZ. In contrast, higher modern [NO] was observed in the Sea of Okhotsk and the Mexican Margin, suggesting that these two study areas have higher oxygen.
All data generated or analyzed during this study are available in the Supplement.
The supplement related to this article is available online at https://doi.org/10.5194/cp-21-1853-2025-supplement.
A.G.M wrote the core manuscript, did the sample preparation, electron microscopy, image and statistical analyses of all the fossil foraminifera. N.G. planned the study design and sampling strategy. G.S. hosted the research group, and provided access to SEM, and lab facilities at the University of Hamburg. D.N. provided sampling material for cores MD01-2415 and M77/2-59-01. C.D. provided sampling material for core MAZ-1E-04. N.L., R.S., and A.B. facilitated the measurement of nitrogen isotopes in the sediment samples of core MD01-2415 and DSDP-480. R.A. contributed to the age model development of core MAZ-1E-04, and H.F helped in the image processing of core DSDP-480. All authors contributed to the discussion and preparation of the manuscript.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We are grateful to the micropaleontology group at the University of Hamburg, Germany. We thank Alfredo Martinez-Garcia, Max Planck Institute for Chemistry, Mainz, Germany for the support with the measurement of nitrogen isotopes in the sediment samples. We acknowledge the help of Jutta Richarz, Kaya Oda for lab support, PhD student Sven Brömme (Max Planck Institute for Chemistry, Mainz) and student assistant Hannah Krüger. We thank Yvon Balut, Agnes Baltzer, and the Shipboard Scientific Party of RV Marion Dufresne cruise WEPAMA 2001 for their kind support. We thank IODP for providing the sample for core DSDP-480. The study is a contribution to the Center for Earth System Research and Sustainability (CEN) of University of Hamburg.
Funding was provided by the Deutsche Forschungsgemeinschaft (DFG) through both N.G.'s Heisenberg grant GL 999/3-1 and grant GL 999/4-1. Funding for the core MD01-2415 recovery was provided by the German Science Foundation (DFG) within project Ti240/11-1. The recovery of core M77-59 recovery was a contribution of the German Science Foundation (DFG) Collaborative Research Project “Climate–Biogeochemistry interactions in the Tropical Ocean” (SFB 754).
This paper was edited by Luc Beaufort and reviewed by two anonymous referees.
Alcorn, R. C., Ontiveros-Cuadras, J. F., Menon, A. G., Glock, N., Tappa, E. J., Burke, J., Cartapanis, O., and Davis, C. V.: Nonlinear Expansion of the Eastern Tropical North Pacific Oxygen Minimum Zone During the Last Deglaciation, Authorea Preprints, https://doi.org/10.22541/essoar.173765742.20874664, 2025.
Altabet, M. A. and Francois, R.: Sedimentary nitrogen isotopic ratio as a recorder for surface ocean nitrate utilization, Global Biogeochemical Cycles, 8, 103–116, https://doi.org/10.1029/93GB03396, 1994.
Altabet, M. A., Pilskaln, C., Thunell, R., Pride, C., Sigman, D., Chavez, F., and Francois, R.: The nitrogen isotope biogeochemistry of sinking particles from the margin of the Eastern North Pacific, Deep Sea Research Part I: Oceanographic Research Papers, 46, 655–679, https://doi.org/10.1016/S0967-0637(98)00084-3, 1999.
Auderset, A., Moretti, S., Taphorn, B., Ebner, P. R., Kast, E., Wang, X. T., Schiebel, R., Sigman, D. M., Haug, G. H., and Martínez-García, A.: Enhanced Ocean oxygenation during Cenozoic warm periods, Nature, 609, 77–82, https://doi.org/10.1038/s41586-022-05017-0, 2022.
Benson, L., Lund, S., Negrini, R., Linsley, B., and Zic, M.: Response of north American Great basin lakes to Dansgaard–Oeschger oscillations, Quaternary Science Reviews, 22, 2239–2251, https://doi.org/10.1016/S0277-3791(03)00210-5, 2003.
Bernhard, J. M., Edgcomb, V. P., Casciotti, K. L., McIlvin, M. R., and Beaudoin, D. J.: Denitrification likely catalyzed by endobionts in an allogromiid foraminifer, The ISME Journal, 6, 951–960, https://doi.org/10.1038/ismej.2011.171, 2012a.
Bernhard, J. M., Casciotti, K. L., McIlvin, M. R., Beaudoin, D. J., Visscher, P. T., and Edgcomb, V. P.: Potential importance of physiologically diverse benthic foraminifera in sedimentary nitrate storage and respiration, J. Geophys. Res., 117, https://doi.org/10.1029/2012JG001949, 2012b.
Bohlen, L., Dale, A. W., and Wallmann, K.. Simple transfer functions for calculating benthic fixed nitrogen losses and C: N: P regeneration ratios in global biogeochemical models, Global Biogeochemical Cycles, 26, https://doi.org/10.1029/2011GB004198, 2012.
Broecker, W. S., Peteet, D. M., and Rind, D.: Does the ocean–atmosphere system have more than one stable mode of operation?, Nature, 315, 21–26, https://doi.org/10.1038/315021a0, 1985.
Bubenshchikova, N., Nürnberg, D., and Tiedemann, R. Variations of Okhotsk Sea oxygen minimum zone: Comparison of foraminiferal and sedimentological records for latest MIS 12–11c and latest MIS 2–1, Marine Micropaleontology, 121, 52–69, https://doi.org/10.1016/j.marmicro.2015.09.004, 2015.
Burke, A. and Robinson, L. F.: The Southern Ocean's role in carbon exchange during the last deglaciation, Science, 335, 557–561, https://doi.org/10.1126/science.1208163, 2012.
Cartapanis, O., Tachikawa, K., and Bard, E.: Northeastern Pacific oxygen minimum zone variability over the past 70 kyr: Impact of biological production and oceanic ventilation, Paleoceanography, 26, https://doi.org/10.1029/2011PA002126, 2011.
Casciotti, K. L.: Nitrite isotopes as tracers of marine N cycle processes, Phil. Trans. R. Soc. A., 374, 20150295, https://doi.org/10.1098/rsta.2015.0295, 2016.
Chang, A. S., Pedersen, T. F., and Hendy, I. L.: Effects of productivity, glaciation, and ventilation on late Quaternary sedimentary redox and trace element accumulation on the Vancouver Island margin, western Canada, Paleoceanography, 29, 730–746, https://doi.org/10.1002/2013PA002581, 2014.
Chang, A. S., Pichevin, L., Pedersen, T. F., Gray, V., and Ganeshram, R.: New insights into productivity and redox-controlled trace element (Ag, Cd, Re, and Mo) accumulation in a 55 kyr long sediment record from Guaymas Basin, Gulf of California, Paleoceanography, 30, 77–94, https://doi.org/10.1002/2014PA002681, 2015.
Cheshire, H. and Thurow, J.: High-resolution migration history of the Subtropical High/Trade Wind system of the northeastern Pacific during the last ∼55 years: Implications for glacial atmospheric reorganization, Paleoceanography, 28, 319–333, https://doi.org/10.1002/palo.20031, 2013.
Choquel, C., Geslin, E., Metzger, E., Filipsson, H. L., Risgaard-Petersen, N., Launeau, P., Giraud, M., Jauffrais, T., Jesus, B., and Mouret, A.: Denitrification by benthic foraminifera and their contribution to N-loss from a fjord environment, Biogeosciences, 18, 327–341, https://doi.org/10.5194/bg-18-327-2021, 2021.
Clark, P., Pisias, N., Stocker, T., and Weaver, A. J.: The role of the thermohaline circulation in abrupt climate change, Nature, 415, 863–869, https://doi.org/10.1038/415863a, 2002.
Clark, P. U. and Mix, A. C.: Ice sheets and sea level of the Last Glacial Maximum, Quaternary Science Reviews, 21, 1–7, https://doi.org/10.1016/S0277-3791(01)00118-4, 2002.
Codispoti, L. A., Brandes, J. A., Christensen, J. P., Devol, A. H., Naqvi, S. W. A., Paerl, H. W., and Yoshinari, T.: The oceanic fixed nitrogen and nitrous oxide budgets: Moving targets as we enter the anthropocene?, Sci. Mar., 65, 85–105, https://doi.org/10.17615/ksfx-e447, 2001.
Dale, A. W., Sommer, S., Lomnitz, U., Bourbonnais, A., and Wallmann, K.: Biological nitrate transport in sediments on the Peruvian margin mitigates benthic sulfide emissions and drives pelagic N loss during stagnation events, Deep Res. I Oceanogr. Res. Pap., 112, 123–136, https://doi.org/10.1016/j.dsr.2016.02.013, 2016.
Devol, A. H., Uhlenhopp, A. G., Naqvi, S. W. A., Brandes, J. A., Jayakumar, D. A., Naik, H., Gaurin, S., Codispoti, L. A., and Yoshinari, T.: Denitrification rates and excess nitrogen gas concentrations in the Arabian Sea oxygen deficient zone, Deep Sea Research Part I: Oceanographic Research Papers, 53, 1533–1547, https://doi.org/10.1016/j.dsr.2006.07.005, 2006.
Dubois, N., Kienast, M., Kienast, S., Normandeau, C., Calvert, S. E., Herbert, T. D., and Mix, A.: Millennial-scale variations in hydrography and biogeochemistry in the Eastern Equatorial Pacific over the last 100 kyr, Quaternary Science Reviews, 30, 210–223, https://doi.org/10.1016/j.quascirev.2010.10.012, 2011.
Dubois, N., Kienast, M., Kienast, S. S., and Timmermann, A.: Millennial-scale Atlantic/East Pacific Sea surface temperature linkages during the last 100,000 years, Earth and Planetary Science Letters, 396, 134–142, https://doi.org/10.1016/j.epsl.2014.04.008, 2014.
Duplessy, J. C., Shackleton, N. J., Fairbanks, R. G., Labeyrie, L., Oppo, D., and Kallel, N.: Deepwater source variations during the last climatic cycle and their impact on the global deepwater circulation, Paleoceanography, 3, 343–360, https://doi.org/10.1029/PA003i003p00343, 1988.
Emmer, E. and Thunell, R. C.: Nitrogen isotope variations in Santa Barbara Basin sediments: Implications for denitrification in the eastern tropical North Pacific during the last 50,000 years, Paleoceanography, 15, 377–387, https://doi.org/10.1029/1999PA000417, 2000.
Erdem, Z., Schönfeld, J., Rathburn, A. E., Pérez, M.-E., Cardich, J., and Glock, N.: Bottom-water deoxygenation at the Peruvian margin during the last deglaciation recorded by benthic foraminifera, Biogeosciences, 17, 3165–3182, https://doi.org/10.5194/bg-17-3165-2020, 2020.
Evans, N., Tichota, J., Ruef, W., Moffett, J., and Devol, A.: Rapid Expansion of Fixed Nitrogen Deficit in the Eastern Pacific Ocean Revealed by 50-Year Time Series, Global Biogeochemical Cycles, 37, e2022GB007575, https://doi.org/10.1029/2022GB007575, 2023.
Fontanier, C., Duros, P., Toyofuku, T., Oguri, K., Koho, K. A., Buscail, R., Grémare, A., Radakovitch, O., Deflandre, B., De Nooijer, L. J., Bichon, S., Goubet, S., Ivanovsky, A., Chabaud, G., Menniti, C., Reichart, G.-J., and Kitazato, H.: Living (stained) deep-sea foraminifera off hachinohe (NE Japan, western Pacific): environmental interplay in oxygen-depleted ecosystems, J. Foraminifer. Res., 44, 281–299, https://doi.org/10.2113/gsjfr.44.3.281, 2014.
Frölicher, T. L., Aschwanden, M. T., Gruber, N., Jaccard, S. L., Dunne, J. P., and Paynter, D.: Contrasting upper and deep ocean oxygen response to protracted global warming, Global Biogeochemical Cycles, 34, e2020GB006601, https://doi.org/10.1029/2020GB006601, 2020.
Fu, W., Primeau, F., Keith Moore, J., Lindsay, K., and Randerson, J. T.: Reversal of increasing tropical ocean hypoxia trends with sustained climate warming, Global Biogeochemical Cycles, 32, 551–564, https://doi.org/10.1002/2017GB005788, 2018.
Galbraith, E. D., Kienast, M., Pedersen, T. F., and Calvert, S. E.: Glacial-interglacial modulation of the marine nitrogen cycle by high-latitude O2 supply to the global thermocline, Paleoceanography, 19, https://doi.org/10.1029/2003PA001000, 2004.
Ganeshram, R. S. and Pedersen, T. F.: Glacial-interglacial variability in upwelling and bioproductivity off NW Mexico: Implications for quaternary paleoclimate, Paleoceanography, 13, 634–645, https://doi.org/10.1029/98PA02508, 1998.
Ganeshram, R. S., Pedersen, T. F., Calvert, S. E., and Murray, J. W.: Large changes in oceanic nutrient inventories from glacial to interglacial periods, Nature, 376, 755–758, https://doi.org/10.1038/376755a0, 1995.
Ganeshram, R. S., Pedersen, T. F., Calvert, S. E., McNeill, G. W., and Fontugne, M. R.: Glacial-interglacial variability in denitrification in the world's oceans: Causes and consequences, Paleoceanography, 15, 361–376, https://doi.org/10.1029/1999PA000422, 2000.
Garcia, H. E., Boyer, T. P., Baranova, O. K., Locarnini, R. A., Mishonov, A. V., Grodsky, A., Paver, C. R., Weathers, K. W., Smolyar, I. V., Reagan, J. R., Seidov, D., and Zweng, M. M.: World Ocean Atlas 2018: product documentation, A. Mishonov, Technical Editor, NOAA Atlas NESDIS, https://doi.org/10.25923/tzyw-rp36, 2019.
Glock, N., Eisenhauer, A., Milker, Y., Liebetrau, V., Schönfeld, J., Mallon, J., Sommer, S., and Hensen, C.: Environmental influences on the pore density of Bolivina spissa (Cushman), J. Foraminifer. Res., 41, 22–32, https://doi.org/10.2113/gsjfr.41.1.22, 2011.
Glock, N., Schönfeld, J., Eisenhauer, A., Hensen, C., Mallon, J., and Sommer, S.: The role of benthic foraminifera in the benthic nitrogen cycle of the Peruvian oxygen minimum zone, Biogeosciences, 10, 4767–4783, https://doi.org/10.5194/bg-10-4767-2013, 2013.
Glock, N., Erdem, Z., Wallmann, K., Somes, C. J., Liebetrau, V., Schönfeld, J., Gorb, S., and Eisenhauer, A.: Coupling of oceanic carbon and nitrogen facilitates spatially resolved quantitative reconstruction of nitrate inventories, Nat. Commun., 9, 1217, https://doi.org/10.1038/s41467-018-03647-5, 2018.
Glock, N., Roy, A., Romero, D., Wein, T., Weissenbach, J., Revsbech, N. P., Høgslund, S., Clemens, D., Sommer, S., and Dagan, T.: Metabolic preference of nitrate over oxygen as an electron acceptor in foraminifera from the Peruvian oxygen minimum zone, Proc. Natl. Acad. Sci. USA, 116, 2860–2865, https://doi.org/10.1073/pnas.1813887116, 2019.
Glock, N., Erdem, Z., and Schönfeld, J.: The Peruvian oxygen minimum zone was similar in extent but weaker during the Last Glacial Maximum than Late Holocene, Communications Earth & Environment, 3, 307, https://doi.org/10.1038/s43247-022-00635-y, 2022.
Gomaa, F., Utter, D. R., Powers, C., Beaudoin, D. J., Edgcomb, V. P., Filipsson, H. L., Hansel, C. M., Wankel, S. D., Zhang, Y., and Bernhard, J. M.: Multiple integrated metabolic strategies allow foraminiferan protists to thrive in anoxic marine sediments, Science Advances, 7, eabf1586, https://doi.org/10.1126/sciadv.abf1586, 2021.
Gorbarenko, S. A., Artemova, A. V., Goldberg, E. L., and Vasilenko, Y. P.: The response of the Okhotsk Sea environment to the orbital-millennium global climate changes during the Last Glacial Maximum, deglaciation and Holocene, Global and Planetary Change, 116, 76–90, https://doi.org/10.1016/j.gloplacha.2014.02.002, 2014.
Govindankutty Menon, A., Davis, C.V., Nürnberg, D., Nomaki, H., Salonen, I., Schmiedl, G., and Glock, N.: A deep-learning automated image recognition method for measuring pore patterns in closely related bolivinids and calibration for quantitative nitrate paleo-reconstructions, Sci. Rep., 13, 19628, https://doi.org/10.1038/s41598-023-46605-y, 2023.
Gray, W. R., Wills, R. C., Rae, J. W., Burke, A., Ivanovic, R. F., Roberts, W. H., Ferreira, D., and Valdes, P. J.: Wind-driven evolution of the North Pacific subpolar gyre over the last deglaciation, Geophysical Research Letters, 47, e2019GL086328, https://doi.org/10.1029/2019GL086328, 2020.
Gruber, N.: The marine nitrogen cycle: overview and challenges, in: Nitrogen in the marine environment, edited by: Capone, D. G., Bronk, D. A., Mulholland, M. R., and Carpenter, E. J., 2nd edn., 1–50, Elsevier, https://doi.org/10.1016/B978-0-12-372522-6.00001-3, 2008.
Hendy, I. L., Pedersen, T. F., Kennett, J. P., and Tada, R.: Intermittent existence of a southern Californian upwelling cell during submillennial climate change of the last 60 kyr, Paleoceanography, 19, https://doi.org/10.1029/2003PA000965, 2004.
Herguera, J. C., Herbert, T., Kashgarian, M., and Charles, C.: Intermediate and deep-water mass distribution in the Pacific during the Last Glacial Maximum inferred from oxygen and carbon stable isotopes, Quaternary Sci. Rev., 29, 1228–1245, https://doi.org/10.1016/j.quascirev.2010.02.009, 2010.
Hess, A. V., Auderset, A., Rosenthal, Y., Miller, K. G., Zhou, X., Sigman, D. M., and Martínez-García, A.: A well-oxygenated eastern tropical Pacific during the warm Miocene, Nature, 619, 521–525, https://doi.org/10.1038/s41586-023-06104-6, 2023.
Holbourn, A., Kiefer, T., Pflaumann, U., and Rothe, S.: WEPAMA Cruise MD 122/IMAGES VII, Rapp. Campagnes Mer OCE/2002/01, Inst. Polaire Fr. Paul Emile Victor (IPEV), Plouzane, France, 2002.
Jaccard, S. and Galbraith, E.: Large climate-driven changes of oceanic oxygen concentrations during the last deglaciation, Nature Geosci., 5, 151–156, https://doi.org/10.1038/ngeo1352, 2012.
Kamp, A., Høgslund, S., Risgaard-Petersen, N., and Stief, P.: Nitrate storage and dissimilatory nitrate reduction by eukaryotic microbes, Front. Microbiol., 6, 1492, https://doi.org/10.3389/fmicb.2015.01492, 2015.
Karstensen, J. and Quadfasel, D.: Formation of Southern Hemisphere thermocline waters: water mass conversion and subduction, J. Phys. Oceanogr., 32, 3020–3038, https://doi.org/10.1175/1520-0485(2002)032<3020:FOSHTW>2.0.CO;2, 2002.
Keeling, R. F., Kortzinger, A., and Gruber, N.: Ocean deoxygenation in a warming world, Ann. Rev.Mar. Sci., 2, 199–229, https://doi.org/10.1146/annurev.marine.010908.163855, 2010.
Keigwin, L. D.: Glacial-age hydrography of the far northwest Pacific Ocean, Paleoceanography, 13, 323–339, https://doi.org/10.1029/98PA00874, 1998.
Keigwin, L. D. and Jones, G. A.: Deglacial climatic oscillations in the Gulf of California, Paleoceanography, 5, 1009–1023, https://doi.org/10.1029/PA005i006p01009, 1990.
Kienast, M., Higginson, M. J., Mollenhauer, G., Eglinton, T. I., Chen, M. T., and Calvert, S. E.: On the sedimentological origin of down-core variations of bulk sedimentary nitrogen isotope ratios, Paleoceanography, 20, https://doi.org/10.1029/2004PA001081, 2005.
Kienast, S. S., Calvert, S. E., and Pedersen, T. F.: Nitrogen isotope and productivity variations along the northeast Pacific margin over the last 120 kyr: Surface and subsurface paleoceanography, Paleoceanography, 17, 7–1, https://doi.org/10.1029/2001PA000650, 2002.
Koutavas, A., Demenocal, P. B., Olive, G. C., and Lynch-Stieglitz, J.: Mid-Holocene El Niño-Southern Oscillation (ENSO) attenuation revealed by individual foraminifera in eastern tropical Pacific sediments, Geology, 34, 993–996, https://doi.org/10.1130/G22810A.1, 2006.
Kuhlmann, H., Freudenthal, T., Helmke, P., and Meggers, H.: Reconstruction of paleoceanography off NW Africa during the last 40,000 years: influence of local and regional factors on sediment accumulation, Marine Geology, 207, 209–224, https://doi.org/10.1016/j.margeo.2004.03.017, 2004.
Kutzbach, J. E. and Wright Jr., H. E.: Simulation of the climate of 18,000 years BP: Results for the North American/North Atlantic/European sector and comparison with the geologic record of North America, Quaternary Science Reviews, 4, 147–187, https://doi.org/10.1016/0277-3791(85)90024-1, 1985.
Lam, P. and Kuypers, M. M. M.: Microbial nitrogen cycling processes in oxygen minimum zones, Ann. Rev. Mar. Sci., 3, 317–345, https://doi.org/10.1146/annurev-marine-120709-142814, 2011.
Leclaire, J. and Kerry, K.: Calcium carbonate and organic carbon stratigraphy of late Quaternary laminated and homogeneous diatom oozes from the Guaymas Slope, HPC Site 480, Gulf of California, in: Initial Reports of the Deep Sea Drilling Project, Vol. 64, edited by: Curray, J. R., Moore, D. G., and the Leg 64 Science Party, U.S. Government Printing Office, Washington, DC, 1263–1275, https://doi.org/10.2973/dsdp.proc.64.168.1982, 1982.
Lembke-Jene, L., Tiedemann, R., Nürnberg, D., Gong, X., and Lohmann, G.: Rapid shift and millennial-scale variations in Holocene North Pacific Intermediate Water ventilation, Proceedings of the National Academy of Sciences, 115, 5365–5370, https://doi.org/10.1073/pnas.1714754115, 2018.
Levin, L. A.: Oxygen minimum zone benthos: Adaptation and community response to hypoxia, in: Oceanography and Marine Biology: An Annual Review, Vol. 41, edited by: Gibson, R. N. and Atkinson, J. A., CRC Press, Boca Raton, FL, 1–45, https://doi.org/10.1201/9780203180570-3, 2003.
Levin, L. A.: Manifestation, drivers, and emergence of open ocean deoxygenation, Annual Review of Marine Science, 10, 229–260, https://doi.org/10.1146/annurev-marine-121916-063359, 2018.
Li, N., Somes, C. J., Landolfi, A., Chien, C.-T., Pahlow, M., and Oschlies, A.: Global impact of benthic denitrification on marine N2 fixation and primary production simulated by a variable-stoichiometry Earth system model, Biogeosciences, 21, 4361–4380, https://doi.org/10.5194/bg-21-4361-2024, 2024.
Liu, K. K. and Kaplan, I. R.: The eastern tropical Pacific as a source of 15N-enriched nitrate in seawater off southern California, Limnology and Oceanography, 34, 820–830, https://doi.org/10.4319/lo.1989.34.5.0820, 1989.
Lourey, M. J., Trull, T. W., and Sigman, D. M.: Sensitivity of δ15N of nitrate, surface suspended and deep sinking particulate nitrogen to seasonal nitrate depletion in the Southern Ocean, Global Biogeochemical Cycles, 17, https://doi.org/10.1029/2002GB001973, 2003.
Martinez, P. and Robinson, R. S.: Increase in water column denitrification during the last deglaciation: the influence of oxygen demand in the eastern equatorial Pacific, Biogeosciences, 7, 1–9, https://doi.org/10.5194/bg-7-1-2010, 2010.
Meckler, A. N., Ren, H., Sigman, D. M., Gruber, N., Plessen, B., Schubert, C. J., and Haug, G. H.: Deglacial nitrogen isotope changes in the Gulf of Mexico: Evidence from bulk sedimentary and foraminifera-bound nitrogen in Orca Basin sediments, Paleoceanography, 26, https://doi.org/10.1029/2011PA002156, 2011.
Meissner, K. J., Galbraith, E. D., and Völker, C.: Denitrification under glacial and interglacial conditions: A physical approach, Paleoceanography, 20, https://doi.org/10.1029/2004PA001083, 2005.
Mollenhauer, G., Schneider, R. R., Müller, P. J., Spieß, V., and Wefer, G.: Glacial/interglacial variablity in the Benguela upwelling system: Spatial distribution and budgets of organic carbon accumulation, Global Biogeochemical Cycles, 16, 81-1, https://doi.org/10.1029/2001GB001488, 2002.
Mollier-Vogel, E., Leduc, G., Böschen, T., Martinez, P., and Schneider, R. R.: Rainfall response to orbital and millennial forcing in northern Peru over the last 18 ka, Quaternary Science Reviews, 76, 29–38, https://doi.org/10.1016/j.quascirev.2013.06.021, 2013.
Mollier-Vogel, E., Martinez, P., Blanz, T., Robinson, R., Desprat, S., Etourneau, J., Charlier, K., and Schneider, R. R.: Mid-holocene deepening of the Southeast Pacific oxycline, Glob. Planet. Change, 172, 365–373, https://doi.org/10.1016/j.gloplacha.2018.10.020, 2019.
Montoya, J. P., Horrigan, S. G., and McCarthy, J. J.: Natural abundance of 15N in particulate nitrogen and zooplankton in the Chesapeake Bay, Marine Ecology Progress Series, 35–61, https://www.jstor.org/stable/24844633, 1990.
Moore, C. M., Mills, M. M., Arrigo, K. R., Berman-Frank, I., Bopp, L., Boyd, P. W., Galbraith, E. D., Geider, R. J., Guieu, C., Jaccard, S. L., and Jickells, T. D.: Processes and patterns of oceanic nutrient limitation, Nat. Geosci., 6, 701–710, https://doi.org/10.1038/ngeo1765, 2013.
Moretti, S., Auderset, A., Deutsch, C., Schmitz, R., Gerber, L., Thomas, E., Luciani, V., Petrizzo, M. R., Schiebel, R., Tripati, A., Sexton, P., Norris, R., D'Onofrio, R., Zachos, J., Sigman, D. M., Haug, G. H., and Martínez-García, A.: Oxygen rise in the tropical upper ocean during the Paleocene-Eocene Thermal Maximum. Science, 383, 727–731, https://doi.org/10.1126/science.adh4893, 2024.
Muratli, J. M., Chase, Z., Mix, A. C., and McManus, J.: Increased glacial-age ventilation of the Chilean margin by Antarctic Intermediate Water, Nature Geoscience, 3, 23–26, https://doi.org/10.1038/ngeo715, 2010.
Naafs, B. D. A., Monteiro, F. M., Pearson, A., Higgins, M. B., Pancost, R. D., and Ridgwell, A.: Fundamentally different global marine nitrogen cycling in response to severe ocean deoxygenation, Proceedings of the National Academy of Sciences, 116, 24979–24984, https://doi.org/10.1073/pnas.1905553116, 2019.
Narita, H., Sato, M., Tsunogai, S., Murayama, M., Ikehara, M., Nakatsuka, T., Wakatsuchi, M., Harada, N., and Ujiié, Y.: Biogenic opal indicating less productive northwestern North Pacific during the glacial ages, Geophysical Research Letters, 29, 22–1, https://doi.org/10.1029/2001GL014320, 2002.
Nürnberg, D., Böschen, T., Doering, K., Mollier-Vogel, E., Raddatz, J., and Schneider, R.: Sea surface and subsurface circulation dynamics off equatorial Peru during the last ∼17 kyr, Paleoceanography, 30, 984–999, https://doi.org/10.1002/2014PA002706, 2015.
Nürnberg, D. and Tiedemann, R.: Environmental change in the Sea of Okhotsk during the last 1.1 million years, Paleoceanography 19, 1–23, https://doi.org/10.1029/2004PA001023, 2004.
Ohkushi, K., Kennett, J. P., Zeleski, C. M., Moffitt, S. E., Hill, T. M., Robert, C., Beaufort, L., and Behl, R. J.: Quantified intermediate water oxygenation history of the NE Pacific: A new benthic foraminiferal record from Santa Barbara basin, Paleoceanography, 28, 453–467, https://doi.org/10.1002/palo.20043, 2013.
Okazaki, Y., Timmermann, A., Menviel, L., Harada, N., Abe-Ouchi, A., Chikamoto, M. O., Mouchet, A., and Asahi, H.: Deepwater formation in the North Pacific during the last glacial termination, Science, 329, 200–204, https://doi.org/10.1126/science.1190612, 2010.
Orsi, W. D., Morard, R., Vuillemin, A., Eitel, M., Wörheide, G., Milucka, J., and Kucera, M.: Anaerobic metabolism of Foraminifera thriving below the seafloor, The ISME Journal, 14, 2580–2594, https://doi.org/10.1038/s41396-020-0708-1, 2020.
Oschlies, A.: A committed fourfold increase in ocean oxygen loss, Nat. Commun., 12, 2307, https://doi.org/10.1038/s41467-021-22584-4, 2021.
Pennington, J. T., Mahoney, K. L., Kuwahara, V. S., Kolber, D. D., Calienes, R., and Chavez, F. P.: Primary production in the eastern tropical Pacific: A review, Progress in oceanography, 69, 285–317, https://doi.org/10.1016/j.pocean.2006.03.012, 2006.
Piña-Ochoa, E., Høgslund, S., Geslin, E., Cedhagen, T., Revsbech, N. P., Nielsen, L. P., Schweizer, M., Jorissen, F., Rysgaard, S., and Risgaard-Petersen, N.: Widespread occurrence of nitrate storage and denitrification among Foraminifera and Gromiida, Proc. Natl. Acad. Sci. USA, 107, 1148–1153, https://doi.org/10.1073/pnas.0908440107, 2010a.
Piña-Ochoa, E., Koho, K. A., Geslin, E., and Risgaard-Petersen, N.: Survival and life strategy of the foraminiferan Globobulimina turgida through nitrate storage and denitrification, Mar. Ecol. Prog. Ser., 417, 39–49, https://doi.org/10.3354/meps, 2010b.
Pospelova, V., Price, A. M., and Pedersen, T. F.: Palynological evidence for late Quaternary climate and marine primary productivity changes along the California margin, Paleoceanography, 30, 877–894, https://doi.org/10.1002/2014PA002728, 2015.
Pride, C., Thunell, R., Sigman, D., Keigwin, L., Altabet, M., and Tappa, E.: Nitrogen isotopic variations in the Gulf of California since the last deglaciation: Response to global climate change, Paleoceanography, 14, 397–409, https://doi.org/10.1029/1999PA900004, 1999.
Pride, C. J.: An evaluation and application of paleoceanographic proxies in the Gulf of California, PhD dissertation, University of South Carolina, 1997.
Rae, J. W., Gray, W. R., Wills, R. C. J., Eisenman, I., Fitzhugh, B., Fotheringham, M., Shevenell, A. E., Taylor, B., and Burke, A.: Overturning circulation, nutrient limitation, and warming in the Glacial North Pacific, Sci. Adv., 6, eabd1654, https://doi.org/10.1126/sciadv.abd1654, 2020.
Rae, J. W. B., Burke, A., Robinson, L. F., Foster, G. L., and Elliott, T.: CO2 storage and release in the deep Southern Ocean on millennial to centennial timescales, Nature, 562, 569–573, https://doi.org/10.1038/s41586-018-0614-0, 2018.
Rafter, P. A., Gray, W. R., Hines, S. K. V., Burke, A., Costa, K. M., Gottschalk, J., Hain, M. P., Rae, J. W. B., Southon, J. R., Walczak, M. H., Yu, J., Adkins, J. F., and DeVries, T.: Global reorganization of deep-sea circulation and carbon storage after the last ice age, Science Advances, 8, eabq5434, https://doi.org/10.1126/sciadv.abq5434, 2022.
Rakshit, S., Glock, N., Dale, A. W., Armstrong, M. M., Scholz, F., Mutzberg, A., and Algar, C. K.: Foraminiferal denitrification and deep bioirrigation influence benthic biogeochemical cycling in a seasonally hypoxic fjord. Geochim. Cosmochim. Acta, 388, 268–282, https://doi.org/10.1016/j.gca.2024.10.010, 2025.
Ren, H., Sigman, D. M., Chen, M. T., and Kao, S. J.: Elevated foraminifera-bound nitrogen isotopic composition during the last ice age in the South China Sea and its global and regional implications, Global Biogeochem. Cycles, 26, GB1020, https://doi.org/10.1029/2010GB004020, 2012.
Riechelson, H., Rosenthal, Y., Bova, S., and Robinson, R. S.: Southern Ocean biological pump role in driving Holocene atmospheric CO2: Reappraisal, Geophysical Research Letters, 51, e2023GL105569, https://doi.org/10.1029/2023GL105569, 2024.
Risgaard-Petersen, N., Langezaal, A. M., Ingvardsen, S., Schmid, M. C., Jetten, M. S. M., Op den Camp, H. J. M., Derksen, J. W. M., Piña-Ochoa, E., Eriksson, S. P., Nielsen, L. P., Revsbech, N. P., Cedhagen, T., and van der Zwaan, G. J.: Evidence for complete denitrification in a benthic foraminifer, Nature, 443, 93–96, https://doi.org/10.1038/nature05070, 2006.
Robinson, R. S., Sigman, D. M., DiFiore, P. J., Rohde, M. M., Mashiotta, T. A., and Lea, D. W.: Diatom-bound 15N/14N: New support for enhanced nutrient consumption in the ice age subantarctic, Paleoceanography, 20, https://doi.org/10.1029/2004PA001114, 2005.
Robinson, R. S., Mix, A., and Martinez, P.: Southern Ocean control on the extent of denitrification in the southeast Pacific over the last 70 ka, Quaternary Science Reviews, 26, 201–212, https://doi.org/10.1016/j.quascirev.2006.08.005, 2007.
Robinson, R. S., Martinez, P., Pena, L. D., and Cacho, I.: Nitrogen isotopic evidence for deglacial changes in nutrient supply in the eastern equatorial Pacific, Paleoceanography, 24, https://doi.org/10.1029/2008PA001702, 2009.
Romanova, V., Lohmann, G., Grosfeld, K., and Butzin, M.: The relative role of oceanic heat transport and orography on glacial climate, Quaternary Science reviews, 25, 832–845, https://doi.org/10.1016/j.quascirev.2005.07.007, 2006.
Russell, J. L. and Dickson, A. G.: Variability in oxygen and nutrients in South Pacific Antarctic intermediate water, Global Biogeochemical Cycles, 17, https://doi.org/10.1029/2000GB001317, 2003.
Salvatteci, R., Gutiérrez, D., Field, D., Sifeddine, A., Ortlieb, L., Bouloubassi, I., Boussafir, M., Boucher, H., and Cetin, F.: The response of the Peruvian Upwelling Ecosystem to centennial-scale global change during the last two millennia, Clim. Past, 10, 715–731, https://doi.org/10.5194/cp-10-715-2014, 2014.
Salvatteci, R., Gutiérrez, D., Sifeddine, A., Ortlieb, L., Druffel, E., Boussafir, M., and Schneider, R.: Centennial to millennial-scale changes in oxygenation and productivity in the Eastern Tropical South Pacific during the last 25,000 years, Quaternary Science Reviews, 131, 102–117, https://doi.org/10.1016/j.quascirev.2015.10.044, 2016.
Salvatteci, R., Schneider, R. R., Blanz, T., and Mollier-Vogel, E.: Deglacial to Holocene ocean temperatures in the Humboldt Current System as indicated by alkenone paleothermometry, Geophysical Research Letters, 46, 281–292, https://doi.org/10.1029/2018GL080634, 2019.
Schlitzer, R.: Ocean Data View, https://odv.awi.de (last access: 25 October 2023), 2023.
Schmidtko, S., Stramma, L., and Visbeck, M.: Decline in global oceanic oxygen content during the past five decades, Nature, 542, 335–339, https://doi.org/10.1038/nature21399, 2017.
Scholz, F., McManus, J., Mix, A. C., Hensen, C., and Schneider, R. R.: The impact of ocean deoxygenation on iron release from continental margin sediments, Nature Geoscience, 7, 433–437, https://doi.org/10.1038/ngeo2162, 2014.
Scholz, F., Schmidt, M., Hensen, C., Eroglu, S., Geilert, S., Gutjahr, M., and Liebetrau, V.: Shelf-to-basin iron shuttle in the Guaymas Basin, Gulf of California, Geochimica et Cosmochimica Acta, 261, 76–92, https://doi.org/10.1016/j.gca.2019.07.006, 2019.
Schubert, C. J. and Calvert, S. E.: Nitrogen and carbon isotopic composition of marine and terrestrial organic matter in Arctic Ocean sediments: implications for nutrient utilization and organic matter composition, Deep Sea Research Part I: Oceanographic Research Papers, 48, 789–810, https://doi.org/10.1016/S0967-0637(00)00069-8, 2001.
Seki, O., Ikehara, M., Kawamura, K., Nakatsuka, T., Ohnishi, K., Wakatsuchi, M., Narita, T., and Sakamoto, T.: Reconstruction of paleoproductivity in the Sea of Okhotsk over the last 30 kyr, Paleoceanography, 19, https://doi.org/10.1029/2002PA000808, 2004.
Sigman, D. M., Altabet, M. A., Michener, R., McCorkle, D. C., Fry, B., and Holmes, R. M.: Natural abundance-level measurement of the nitrogen isotopic composition of oceanic nitrate: an adaptation of the ammonia diffusion method, Marine Chemistry, 57, 227–242, https://doi.org/10.1016/S0304-4203(97)00009-1, 1997.
Skinner, L. C., Fallon, S., Waelbroeck, C., Michel, E., and Barker, S.: Ventilation of the deep Southern Ocean and deglacial CO2 rise, Science, 328, 1147–1151, https://doi.org/10.1126/science.1183627, 2010.
Somes, C. J., Schmittner, A., Muglia, J., and Oschlies, A.: A three-dimensional model of the marine nitrogen cycle during the last glacial maximum constrained by sedimentary isotopes, Frontiers in Marine Science, 4, 108, https://doi.org/10.3389/fmars.2017.00108, 2017.
Stramma, L., Johnson, G. C., Sprintall, J., and Mohrholz, V.: Expanding oxygen-minimum zones in the tropical oceans, Science, 320, 655–658, https://doi.org/10.1126/science.1153847, 2008.
Stramma, L., Schmidtko, S., Levin, L. A., and Johnson, G. C.: Ocean oxygen minima expansions and their biological impacts, Deep-Sea Res. I Ocean. Res. Pap., 57, 587–595, https://doi.org/10.1016/j.dsr.2010.01.005, 2010.
Studer, A. S., Mekik, F., Ren, H., Hain, M. P., Oleynik, S., Martínez-García, A., Jaccard, S. L., and Sigman, D. M.: Ice age-Holocene similarity of foraminifera-bound nitrogen isotope ratios in the eastern equatorial Pacific, Paleoceanography and Paleoclimatology, 36, e2020PA004063, https://doi.org/10.1029/2020PA004063, 2021.
Takano, Y., Ito, T., and Deutsch, C.: Projected centennial oxygen trends and their attribution to distinct ocean climate forcings, Global Biogeochemical Cycles, 32, 1329–1349, https://doi.org/10.1029/2018GB005939, 2018.
Taylor, M. A., Hendy, I. L., and Chappaz, A.: Assessing oxygen depletion in the Northeastern Pacific Ocean during the last deglaciation using I/Ca ratios from multiple benthic foraminiferal species, Paleoceanography, 32, 746–762, https://doi.org/10.1002/2016PA003062, 2017.
Ternois, Y., Kawamura, K., Keigwin, L., Ohkouchi, N., and Nakatsuka, T.: A biomarker approach for assessing marine and terrigenous inputs to the sediments of Sea of Okhotsk for the last 27,000 years, Geochimica et Cosmochimica Acta, 65, 791–802, https://doi.org/10.1016/S0016-7037(00)00598-6, 2001.
Tesdal, J.-E., Galbraith, E. D., and Kienast, M.: Nitrogen isotopes in bulk marine sediment: linking seafloor observations with subseafloor records, Biogeosciences, 10, 101–118, https://doi.org/10.5194/bg-10-101-2013, 2013.
Thunell, R. C., Sigman, D. M., Muller-Karger, F., Astor, Y., and Varela, R.: Nitrogen isotope dynamics of the Cariaco Basin, Venezuela, Global Biogeochemical Cycles, 18, https://doi.org/10.1029/2003GB002185, 2004.
Ujiié, H. and Ujiié, Y.: Late Quaternary course changes of the Kuroshio Current in the Ryukyu Arc region, northwestern Pacific Ocean, Marine Micropaleontology, 37, 23–40, https://doi.org/10.1016/S0377-8398(99)00010-9, 1999.
Van Geen, A., Fairbanks, R. G., Dartnell, P., McGann, M., Gardner, J. V., and Kashgarian, M.: Ventilation changes in the northeast Pacific during the last deglaciation, Paleoceanography, 11, 519–528, https://doi.org/10.1029/96PA01860, 1996.
Voss, M., Bange, H. W., Dippner, J. W., Middelburg, J. J., Montoya, J. P., and Ward, B.: The marine nitrogen cycle: recent discoveries, uncertainties and the potential relevance of climate change, Philos. Trans. R. Soc. B, 368, 293–296, https://doi.org/10.1098/rstb.2013.0121, 2013.
Wada, E. and Hattori, A.: Nitrogen isotope effects in the assimilation of inorganic nitrogenous compounds by marine diatoms, Geomicrobiol. J., 1, 85–101, https://doi.org/10.1080/01490457809377725, 1978.
Waelbroeck, C., Frank, N., Jouzel, J., Parrenin, F., Masson-Delmotte, V., and Genty, D.: Transferring radiometric dating of the last interglacial sea level high stand to marine and ice core records, Earth Planet. Sci. Lett., 265, 183–194, https://doi.org/10.1016/j.epsl.2007.10.006, 2008.
Wallmann, K., Schneider, B., and Sarnthein, M.: Effects of eustatic sea-level change, ocean dynamics, and nutrient utilization on atmospheric pCO2 and seawater composition over the last 130 000 years: a model study, Clim. Past, 12, 339–375, https://doi.org/10.5194/cp-12-339-2016, 2016.
Wallmann, K., José, Y. S., Hopwood, M. J., Somes, C. J., Dale, A. W., Scholz, F., Grasse, P., and Oschlies, A.: Biogeochemical feedbacks may amplify ongoing and future ocean deoxygenation: a case study from the Peruvian oxygen minimum zone, Biogeochemistry, 159, 45–67, https://doi.org/10.1007/s10533-022-00908-w, 2022.
Wang, Y., Hendy, I. L., and Thunell, R.: Local and remote forcing of denitrification in the northeast Pacific for the last 2,000 years, Paleoceanogr. Paleoclimatol., 34, 1517–1533, https://doi.org/10.1029/2019PA003577, 2019.
Wang, Y., Hendy, I. L., and Zhu, J.: Expansion of the Southern California oxygen minimum zone during the early-to mid-Holocene due to reduced ventilation of the Northeast Pacific, Quaternary Sci. Rev., 238, 106326, https://doi.org/10.1016/j.quascirev.2020.106326, 2020.
White, A. E., Foster, R. A., Benitez-Nelson, C. R., Masqué, P., Verdeny, E., Popp, B. N., Arthur, K. E., and Prahl, F. G.: Nitrogen fixation in the Gulf of California and the eastern tropical North Pacific, Prog. Oceanogr., 109, 1–17, https://doi.org/10.1016/j.pocean.2012.09.002, 2013.
Woehle, C., Roy, A.-S., Glock, N., Wein, T., Weissenbach, J., Rosenstiel, P., Hiebenthal, C., Michels, J., Schönfeld, J., and Dagan, T.: A Novel Eukaryotic Denitrification Pathway in Foraminifera, Curr. Biol., 28, 2536–2543, https://doi.org/10.1016/j.cub.2018.06.027, 2018.
Woehle, C., Roy, A., Glock, N., Michels, J., Wein, T., Weissenbach, J., Romero, D., Hiebenthal, C., Gorb, S. N., Schönfeld, J., and Dagan, T.: Denitrification in foraminifera has an ancient origin and is complemented by associated bacteria, Proc. Natl. Acad. Sci. USA, 119, e2200198119, https://doi.org/10.1073/pnas.2200198119, 2022.
Yamamoto, A., Abe-Ouchi, A., Shigemitsu, M., Oka, A., Takahashi, K., Ohgaito, R., and Yamanaka, Y.: Global deep ocean oxygenation by enhanced ventilation in the Southern Ocean under long-term global warming, Global Biogeochemical Cycles, 29, 1801–1815, https://doi.org/10.1002/2015GB005181, 2015.