the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 23 Oct 2025
| 23 Oct 2025
Late Pleistocene temperature patterns in the Western Palearctic: insights from rodent associations compared with general circulation models
Aurélien Royer
Julien Crétat
Rémi Laffont
Sara Gamboa
Belén Luna
Iris Menéndez
Benjamin Pohl
Sophie Montuire
Manuel Hernández Fernández
Since rodent fossils are preserved in many low- and high-latitude archaeological and paleontological sites from a wide variety of environments, their associations are a commonly useful proxy for inferring past local climate and environmental conditions. Such a frequent and widespread geographic distribution can help us to better understand past climate evolution by providing access to high spatiotemporal resolution at large geographical scales. The aim of this paper is to develop an approach to generate continental-scale temperature maps based on rodent associations and to assess their reliability compared to state-of-the-art general circulation models (GCMs). We used the Bioclimatic Analysis, based on fossil and modern rodent associations, to infer climate zone distribution and local temperatures (mean annual temperature, mean temperature of the warmest month and mean temperature of the coldest month), at the Western Palearctic (Europe, Middle East and North Africa) for six different periods: the Last Glacial Maximum (LGM), Heinrich Stadial, Bølling, Allerød, Younger Dryas and present-day conditions. The Bioclimatic Analysis is combined with a spatial generalized linear mixed model to interpolate these surface temperatures across the Western Palearctic. We show that the spatial patterns in mean annual temperature and mean temperature of the warmest and coldest months are very similar between our interpolations and GCMs for both present-day and LGM conditions, but the rodent-based approach provides slightly cooler LGM estimations in western Europe and warmer ones in eastern Europe. Throughout the Late Glacial oscillations, the rodent-based model infers globally small variations in mean annual temperature and mean temperature of the warmest months and slightly larger changes in mean temperature of the coldest months. Nonetheless, some events show weak but significant regional variations depending of the events and the climate variable. For instance, the most important shifts in mean annual temperature between the Allerød and Younger Dryas are observed in northwestern regions. Northeastern regions, on the other hand, experienced relatively stable mean annual temperature, although they did experience considerable warming of the warmest month and cooling of the coldest month. Minor discrepancies appear between GCMs and the rodent-based model, the latter showing colder temperature in northwestern Europe and hence a differential west–east gradient in ice-sheet influence. Our results demonstrate that rodent associations are robust proxies for reconstructing and regionalizing past temperatures at broad scales, offering a readily reproducible approach to be reimplemented in future studies incorporating new rodent data.
- Article
(18802 KB) - Full-text XML
-
Supplement
(18279 KB) - BibTeX
- EndNote




Understanding past climate changes is necessary for interpreting the evolution of living organisms and their communities in their historical environmental context. Past climate information comes from climate proxy records (in the form of geochemical, physical, geological or biological data) obtained from a natural archive (Markwick, 2007) and/or from general circulation models based on thermodynamic laws (Randall, 2000). Each proxy has its own set of advantages and limitations. Some are directly correlated to climatic variables (e.g. some isotopes), while others reflect their indirect influence (e.g. pollen, invertebrate or vertebrate associations). Nonetheless, all are complementary, because each proxy provides a piece of information about the global system for a specific sphere (e.g. hydrosphere, biosphere), on particular geographical scales and timescales, and at a certain resolution. Paleoclimatic inference methods based on different climate proxies are constantly being improved to overcome their limitations and to provide refined past climate change information. As any single proxy still provides a unique history of the past environment and climate, many studies have emphasized the need for multi-proxy approaches and/or comparisons between data from proxies of different nature (e.g. Allen et al., 2008) to have a better understanding of paleoclimate proxy records and climate variations.
Among the many natural archives (e.g. speleothems, chironomids, pollen) available to approximate past climate changes, fossil remains of small mammals (e.g. Rodentia and/or Eulipotyphla) have been used for a long time because they present several advantages:
-
Their large taxonomic diversity is associated with a wide range of ecological and biological traits that ensure their ubiquitous occurrence in all modern continental biotopes (e.g. Quéré and Le Louarn, 2011; Wilson et al., 2017; Wilson and Mittermeier, 2018).
-
Their small size, associated with high reproductive rates and fast evolutionary change, makes them particularly sensitive to changes in climate and habitat, which constrain their geographical distributions, biotic interactions and, ultimately, community organization (Wolff and Sherman, 2008).
-
They are regularly found in archaeological and paleontological sites (e.g. Hinton, 1926; Chaline, 1972; Daams et al., 1988; Martin et al., 2000; van den Hoek Ostende, 2003; Cuenca-Bescós et al., 2010; Furió et al., 2011; Minwer-Barakat et al., 2012; Erbajeva and Alexeeva, 2013; Royer et al., 2016; Harzhauser et al., 2017; Markova et al., 2019; Flynn et al., 2023).
As a consequence, small vertebrates and in particular rodents have been recognized as a good proxy and are regularly used for inferring past local and continental environmental and climatic conditions (e.g. Simard-Pellissier, 1966; Chaline, 1972; Tchernov, 1975; Avery, 1987; van der Meulen and Daams, 1992; Montuire and Desclaux, 1997; Montuire and Marcolini, 2002; Legendre et al., 2005; Hernández Fernández et al., 2007; García-Alix et al., 2008; Cuenca-Bescós et al., 2009; López-García et al., 2010; Royer et al., 2013, 2014; Rofes et al., 2015; Royer et al., 2016; Fernández-García et al., 2016; Mansino et al., 2016; Jeannet, 2018; Stoetzel et al., 2019; López-García et al., 2021; Royer et al., 2021; Linchamps et al., 2023; García-Morato et al., 2024; Piñero et al., 2024; Schürch et al., 2024).
In parallel, many different fully coupled general circulation models (GCMs) have been developed and used over the past decades to understand past, present-day and future climate variabilities and Earth system changes (e.g. Sloan et al., 1996; Prentice et al., 1998; Salzmann et al., 2008; Haywood et al., 2009; Henrot et al., 2017; Beyer et al., 2020; Kageyama et al., 2021a; Kageyama and Paillard, 2021) and to explore their influence on human communities and environment changes (e.g. Allen et al., 2010; Burke et al., 2021; Leonardi et al., 2023; Albouy et al., 2024). GCMs use the equations of fluid thermodynamics and integrate biotic and abiotic variables to resolve the space–time changes and dynamics of the Earth climate system. GCMs provide numerical climate information at the global scale and are therefore not limited by the uneven distribution of sample locations and taphonomic biases. Comparing results from different climate proxies, such as small mammals, to GCM outputs offers considerable potential for gaining a better understanding of the mechanisms driving climate variability (e.g. Emile-Geay et al., 2016; Carré et al., 2021). It also provides unique opportunities for evaluating model performance and proxy applicability (e.g. Kageyama et al., 2001; Jost et al., 2005; Allen et al., 2010; Latombe et al., 2018; Comas-Bru et al., 2019). Although small mammal associations are generally compared with other proxy data at a local scale, such as pollen or stable isotope compositions, combining paleoclimate inferences based on their assemblages with GCM paleoclimate simulations has been rarely tested for refining environmental and chronological context (e.g. Terray et al., 2023). The integration of such data sources into a general approach to paleoclimatic research yields the potential to attain major advances in the understanding of climate change at large spatial scales.
Rodent fossil remains are abundant at many sites throughout Europe, from the Mediterranean region to high latitudes. This abundance allows us not only to use their assemblages to reconstruct local climate conditions, but also to access continental-scale paleoclimate reconstructions across a wide range of environments and time intervals (e.g. Hernández Fernández, 2006; Puzachenko and Markova, 2019, 2023). In this paper, we use rodent associations to infer surface temperature under different climate conditions. We compare their inferences with GCM outputs to better elucidate faunal community signatures and refine climate reconstructions. We focus on large spatial scales, over the Western Palearctic (Europe, Middle East and North Africa), where a large number of fossil data are available from the Last Glacial Maximum (LGM) to the end of the Late Glacial, from 23 000 to 11 700 years before present. This period of major climate changes is characterized by a succession of short warm (Greenland Interstadial) and cold (Greenland Stadial) events leading to the so-called Bølling, Allerød and Younger Dryas pollen chronozones (e.g. Björck et al., 1998; Peyron et al., 2005; Rasmussen et al., 2014). These climatic events have strongly shaped Palearctic environments and faunal communities.
The aim of this study is twofold:
-
First, we propose a new approach to interpolate surface temperature derived from species composition of rodent associations by combining the Bioclimatic Analysis (Hernández Fernández and Peláez-Campomanes, 2003, 2005; Royer et al., 2020) with a spatial generalized linear mixed model. This approach is used to reconstruct mean annual surface temperature (MAT), as well as mean surface temperature of the warmest (MTWA) and coldest months (MTCO), in the present-day context and for successive events within the LGM–Late Glacial interval.
-
Second, we compare these reconstructions with GCM simulations integrated over the same time periods to determine the degree of spatiotemporal agreement between those approaches.
2.1 Rodent associations from modern localities
A total of 157 modern localities covering northern Africa and western Eurasia (north of 25° N and west of 60° E) were used to reconstruct spatial variation in the present-day climate (Fig. 1; Table S1 in the Supplement). Most of the sites (n=152) are located at low altitude (below 1000 m above current sea level) to focus on large-scale signal with no interference of local topography; only five localities are at an altitude higher than 1000 m above sea level. For each locality, we compiled a rodent faunal list (Table S2) by identifying all rodent species whose geographic range polygons, sourced from the IUCN Red List spatial data (IUCN, 2021), intersect with a 50 km radius buffer around the locality coordinates.
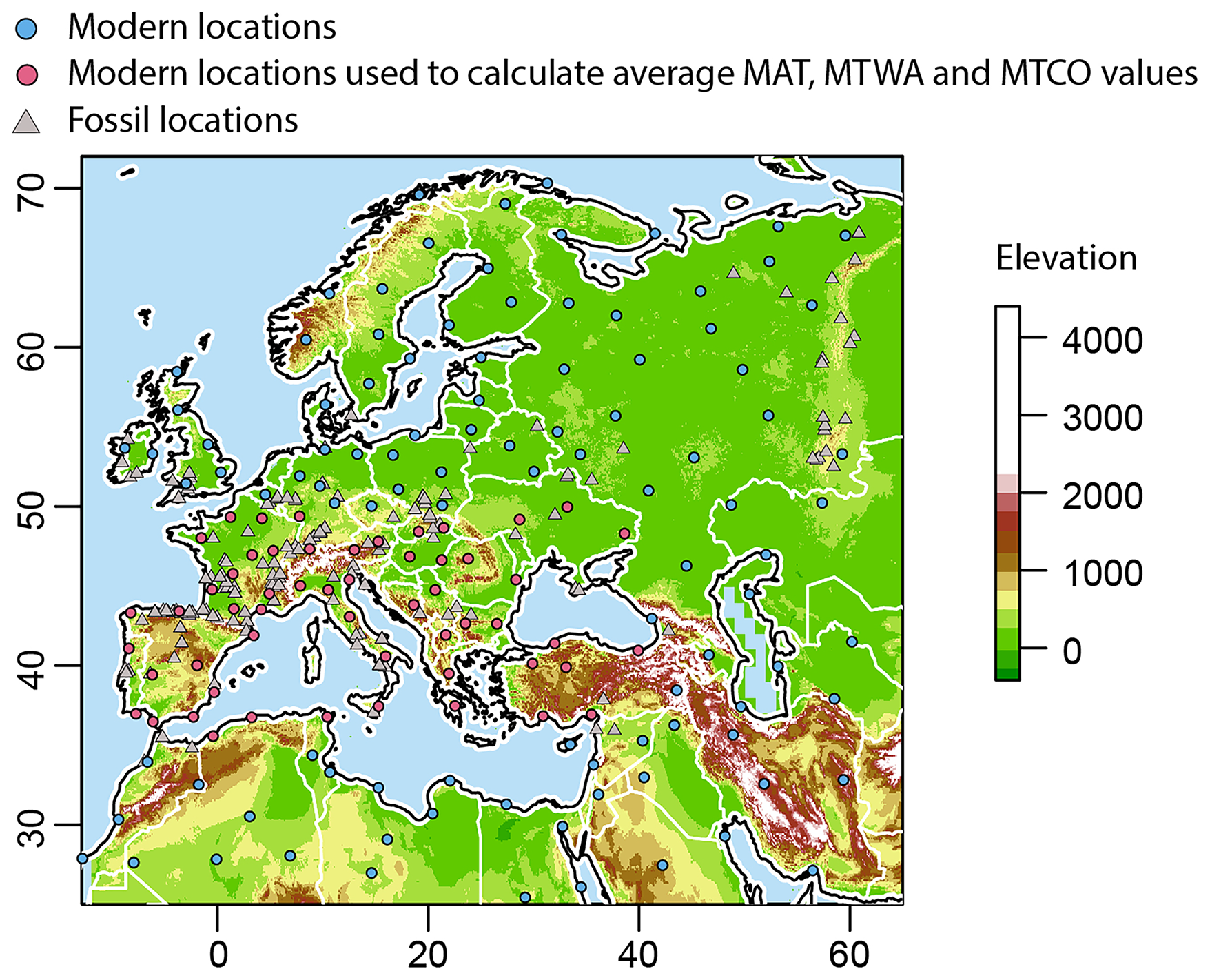
Figure 1Geographical distribution of modern (circles) and fossil (triangles) rodent associations studied in this work. Pink circles show the modern assemblages used to calculate area-averaged MAT, MTWA and MTCO values.
2.2 Temporal framework and rodent associations from fossil localities
We focused on rodent associations from the Last Glacial Maximum (LGM) to the end of Late Glacial, i.e. between 23 000 and 11 700 years before present (BP), compiling a database that includes 279 associations coming from stratigraphic units in 168 archaeological and paleontological sites located from Morocco to the Ural Mountains (Fig. 1, Table S2). These fossil associations do not represent a synchronous ensemble. To maximize fossil data availability, we grouped them into five time intervals that correspond to major climatic events and pollen chronozones: LGM (GS2.1bc: 23 000–18 000 years BP; 65 fossil associations), Heinrich Stadial 1 (GS2.1a: 18 000–14 900 BP; 93 assoc.), Bølling (GI-1de: 14 900–13 900 BP; 47 assoc.), Allerød (GI-1abc: 13 900–12 900 BP; 41 assoc.) and Younger Dryas (GS-1: 12 900–11 700 BP; 33 assoc.).
Reconstructing paleoenvironmental and paleoclimatic conditions from faunal remains requires specific analytical and taphonomical considerations (Lyman, 2019) to ensure high data quality. As such data are derived from biological communities, the quality of faunal data depends on accurate taxonomic identifications. Many species are indeed very difficult to identify due to strong similarity, high morphological variability or extremely low local abundance (e.g. Nadachowski, 1982; Markova et al., 2010; Escudé et al., 2013; Navarro et al., 2018; Arbez et al., 2021; Stoetzel et al., 2023). Faunal remains primarily come from field data, which are regularly re-evaluated to verify the data integrity that may have gone unnoticed during the fieldwork or initial study, in relation to stratigraphic formation, palimpsest processes and excavation unit definition (e.g. Lyman, 2008; Royer, 2014; Discamps et al., 2023), as well as bioturbation (e.g. Pelletier et al., 2017). To select the stratigraphic units, we ensured that the associated rodent assemblages were detailed and representative, maintaining good taphonomical integrity of small mammal remains within the archaeological and paleontological stratigraphic units. Sites that did not meet these criteria were not included, despite their contributions, such as the presence of the collared lemming (Dicrostonyx torquatus) in Croatia (Lenardić, 2013; Lenardić et al., 2018) or the striped field mouse (Apodemus agrarius) in France (Aguilar et al., 2008) during the LGM. Both are found in sites for which unfortunately the associated rodents are unknown. Since each fossil association should be included in a particular time interval, we selected mainly stratigraphic units associated with radiocarbon dates restricted to a single time interval (Table S2). Radiocarbon dates were calibrated at 2σ (95.4 %) using the oxcAAR v1.1.1 R package (Hinz et al., 2018) and IntCal20 calibration curve (Reimer et al., 2020).
2.3 The Bioclimatic Analysis
Many methods have been developed to estimate climatic variables from rodent communities (e.g. Hokr, 1951; van de Weerd and Daams, 1978; Daams and van der Meulen, 1984; Chaline and Brochet, 1989; Sesé, 1991; Montuire et al., 1997; van Dam, 2006; Jeannet, 2010; Fagoaga et al., 2019). We used here the Bioclimatic Analysis approach, originally developed by Hernández Fernández (2001) and Hernández Fernández and Peláez-Campomanes (2003, 2005) and updated by Royer et al. (2020). This approach, detailed in these studies, is a commonly used technique in paleoclimatic studies of fossil sites (e.g. Hernández Fernández, 2006; Hernández Fernández et al., 2007; Pérez-Crespo et al., 2013; Socha, 2014; Laplana et al., 2016; Piñero et al., 2016; López-García et al., 2017; Berto et al., 2019b; Fernández-García et al., 2020; Lemanik et al., 2020; Álvarez-Vena et al., 2021; Izvarin et al., 2022; Jovanović et al., 2022; Luzi et al., 2022b; Arbez et al., 2023; Domínguez-García et al., 2023, 2024; Lebreton and López-García, 2023; Rey-Rodríguez et al., 2024; Stoetzel et al., 2025). It skilfully assesses multiple past climate variables (e.g. temperature and precipitation) from mammal communities and provides reliable estimations as well as their associated uncertainties.
The original models for the Bioclimatic Analysis (Hernández Fernández and Peláez-Campomanes, 2003, 2005) were based on 50 modern localities at global scale and successfully validated using an additional set of different localities from the ones used to develop them. The current models constructed by Royer et al. (2020) were based on a new set of 49 modern communities distributed throughout the Palearctic in order to be representative of the different climate zones (seven localities for seven climate zones). In that paper, the models were also validated based on leave-one-out cross-validation (LOOCV).
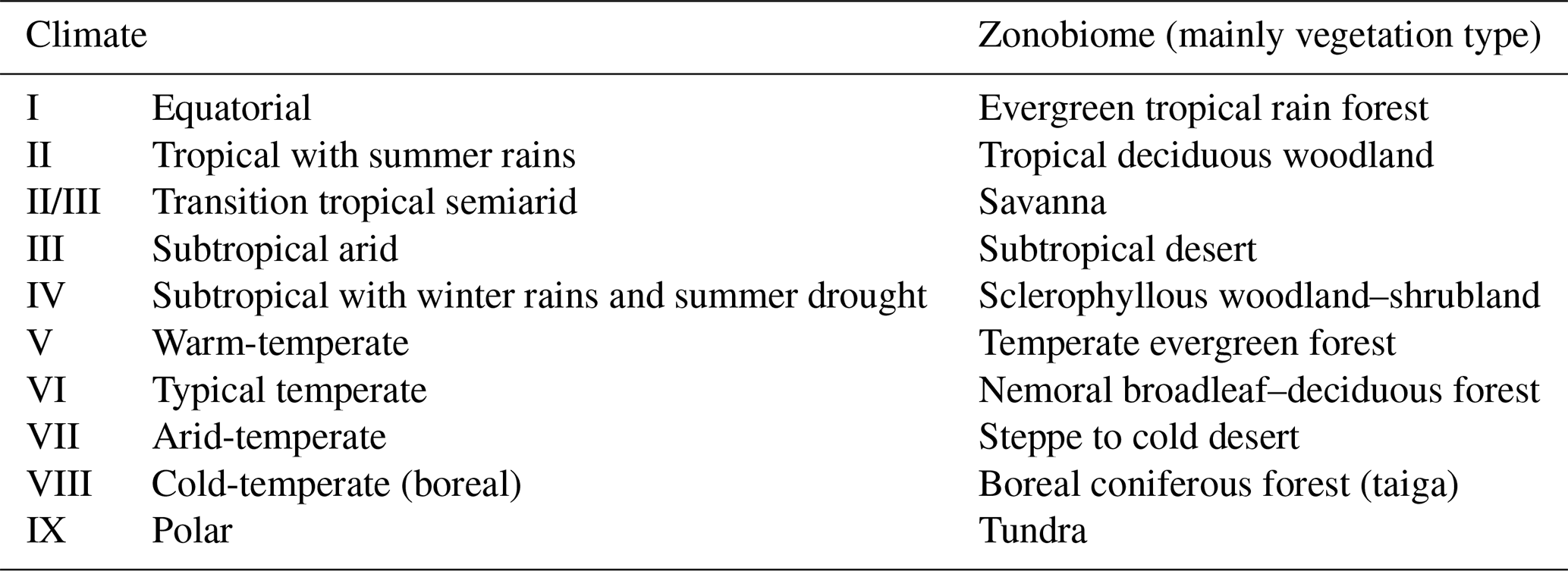
This approach is based on the bioclimatic breadth of each species (assessed through the Climatic Restriction Index, CRI; Hernández Fernández, 2001), which is based on the presence of each species in different climate zones (Table 1), as defined by the combination between monthly precipitation and temperature mean values (Walter, 1970). For this study, we added CRI values for more than 100 species not defined in the original model, including some extinct species and some “chimeric” species, which are taxa only identified at genus levels or not distinguished from morphological sibling species. These CRI values were defined as in the original model, following the recently updated taxonomic and geographical range information (Wilson et al., 2017). All these added CRI values are detailed in Table S3.
Table 1Climate zone typology (from Walter, 1970) and its relationships with world vegetation types.
The bioclimatic spectrum of a locality consists of values of 10 bioclimatic components (BCs; Hernández Fernández, 2001), calculated on the basis of the CRI values for each climate zone across the entire faunal assemblage and detailed in Table S2. The use of BCs helps to address the problem of classifying paleoecological records by reducing the number of entities considered (only 10 BCs) and by providing an ecological basis for treating mammals from different regions in a compatible way.
The Bioclimatic Analysis is composed of two parts. The first one relies on linear discriminant functions deduced from the bioclimatic spectra of modern mammalian communities. These linear discriminant functions are subsequently used to classify additional observations (extinct communities in our case) in each climatic zone, with an associated posterior probability (Hernández Fernández and Peláez-Campomanes, 2003). We used posterior probability values to assess the robustness of the climate classifications obtained by the discriminant functions and considered robust probabilities above 0.95. A prediction error was estimated at around 12 % for estimating the bioclimatic zone with linear discriminant functions based on rodent communities (Royer et al., 2020). The second part of the Bioclimatic Analysis is built from transfer functions by means of multiple linear regression analyses of climatic parameters and modern bioclimatic spectra. Most predictive equations generated by multiple linear regressions for each climatic factor produced highly significant determination coefficients (Hernández Fernández and Peláez-Campomanes, 2005), and rarefaction analysis revealed these new models to be reliable even when a substantial percentage of species from the original community was removed (Royer et al., 2020). These models are ultimately used to infer climatic variables for additional observations (i.e. extinct communities). Although the Bioclimatic Analysis gives the possibility of estimating 11 climatic variables from fossil faunal assemblages (Hernández Fernández and Peláez-Campomanes, 2005; Royer et al., 2020), in this paper, we focus only on 3 of them: the mean annual surface temperature (MAT), the mean surface temperature of the warmest month (MTWA) and the mean surface temperature of the coldest month (MTCO), which are characterized by coefficients of determination of 0.94, 0.92 and 0.85, respectively (Royer et al., 2020).
2.4 Building interpolation maps
Once temperature data for individual localities were calculated, interpolation maps for the whole Western Palearctic at each time interval were built using the “isofit” and “isoscape” functions from the R package IsoriX v.0.9.2 (Courtiol and Rousset, 2017; Courtiol et al., 2019), which is based on the spaMM v.4.4.16 (Rousset et al., 2016), rasterVis v.0.51.6 (Perpinan Lamigueiro et al., 2023) and terra v.1.7 (Hijmans et al., 2022) R packages. Interpolations of the three climatic variables estimated from rodent associations were inferred through a spatial generalized linear mixed model (GLMM), using latitude and longitude as covariates. Although the GLMM can include the effects of altitude, the number of high-altitude fossil sites is too limited per period to obtain sufficiently robust correlations to reproduce the adiabatic effect, leading to erroneous models for geographical areas at middle and high altitudes. We therefore prefer to apply an average vertical drift rate to reflect this effect in geographical and temporal terms. Elevation effects were accounted for by applying a temperature lapse rate of −0.5 °C per 100 m to rodent association Bioclimatic Model results, and by reinjecting it after the spatial interpolation. The original functions of IsoriX were slightly modified to (1) better assess local prediction and uncertainties from the Bioclimatic Model and (2) account for ocean and ice-sheet masks:
-
Initially, the IsoriX package required aggregated data at each locality to directly calculate the mean and variance before building the interpolation maps. We modified the “isoscape” function to use the inferred data and predicted variance derived from applying the Bioclimatic Model instead of aggregated data. The original Bioclimatic Analysis script was also slightly modified to extract the standard error, the residual scale and the degree of freedom of the residuals generated for each locality and to calculate the variance of inferred data used by the isoscape function.
-
In order to obtain approximate boundaries for accessible areas, we generated ocean masks using mean bathymetry variations (Lambeck et al., 2014; Benjamin et al., 2017) of −130 m for the LGM, −120 m for the Heinrich Stadial 1, −100 m for the Bølling, −80 m for the Allerød and −60 m for the Younger Dryas. We also used ice-sheet masks derived from the global ICE-6G_C (VM5a) model (Argus et al., 2014; Peltier et al., 2015), with values at 21.0 kyr for the LGM, 17.0 kyr for the Heinrich Stadial 1, 14.5 kyr for the Bølling, 13.5 kyr for the Allerød and 12.5 kyr for the Younger Dryas. The generated maps have a resolution of 0.1° × 0.1°.
The quality of the models produced and sites used was evaluated using standard deviation calculated from leave-one-out cross-validation (LOOCV) to identify potential outliers to remove (Fig. S1 in the Supplement). Three outliers (Bouârfa, Achgabat and Birjand) were detected and excluded from present-day simulations, and one outlier sizably impacting temperature reconstructions was observed with the Younger Dryas Belarusian site of Volosovo.
2.5 General circulation model simulation of modern and past periods
An essential assessment for any method used in paleoclimatic reconstruction involves comparing inferred climate estimates with observed data. Even though the Bioclimatic Analysis has previously been tested on several occasions (Hernández Fernández and Peláez-Campomanes, 2003, 2005; Royer et al., 2020), we took the opportunity to evaluate its application by including the spatial dimension. To discuss similarities and differences between our rodent-based reconstructions and state-of-the-art GCMs from modern and past climatic conditions, we used ERA5 reanalysis, GCM simulations from the PMIP4 project and GCM simulations published by Beyer et al. (2020).
By combining models and large numbers of observations (e.g. weather stations, satellite and airborne measurements, buoys, radiosonde data), the state-of-the-art ERA5 global reanalysis (Hersbach et al., 2020) provides a spatially and temporally accurate climate dataset at 0.25° resolution, in particular in Europe (e.g. Johannsen et al., 2019). Here, we used ERA5 as the reference for the modern climatic context for assessing the quality of our rodent-based model and GCMs. We have extracted monthly air temperature values 2 m above the surface over land from the 1979–2008 period, from an area bounded by the 25° N latitude and the 55° E longitude (which is therefore slightly less extensive than the rodent model).
The PMIP4 project regroups a large ensemble of GCM simulations built for different time periods, including the current present day and the LGM (Kageyama et al., 2021). For the modern climatic context, we used six PMIP4 simulations for the 1976–2005 period: CCSM4 (Gent et al., 2011), CNRM-CM5 (Voldoire et al., 2013), IPSL-CM5A-LR (Dufresne et al., 2013), MIROC-ESM (Sueyoshi et al., 2013), MPI-ESM-P (Giorgetta et al., 2013) and MRI-CGCM3 (Yukimoto et al., 2012). For the LGM, we used the same six PMIP4 simulations plus the COSMOS-ASO (Budich et al., 2010) and FGOALS-g2 (Zheng and Yu, 2013) simulations. For conciseness, we consider the PMIP4 ensemble mean for spatial mapping.
Beyer et al. (2020) published 2 m air temperature estimations combining medium- and high-resolution GCM (HadCM3 and HadAM3H) simulations and modern-era instrumental data (Beyer et al., 2020). This product, hereafter called Beyer2020, provides monthly bias-corrected 2 m air temperature and many other parameters at the 0.5° spatial resolution and 1000–2000-year temporal resolution for the last 120 000 years, giving the possibility of comparing the different periods of the Late Glacial. We extracted here the Beyer2020 simulations at −21.0, −17.0, −15.0, −13.0, −12.0 and 0 ky BP.
Finally, to quantify global changes between current and past climatic contexts, we selected 55 of the 157 localities as anchors to calculate average MAT, MTWA and MTCO values from our rodent-based reconstructions and the GCMs. These 55 localities cover the 35–50° N/15° W–40° E domain (Fig. 1; Table S1), which was not impacted by ice-sheet development (allowing a comparison between past and present periods) and includes a large part of the archaeological and paleontological sites used in this study, thus limiting the inference extrapolation in data-free areas.
All the above analyses were performed using the R software (R core team, 2023). R scripts and raw data are available in Supplement S1, in order to be fully reproducible.
3.1 A simple approach from a complex system
Caution must always be exercised when working with reconstructions based on fossil faunal associations (e.g. Lyman, 2017). Like any methodological approach based on biotic associations recorded in fossil sites, rodent-based paleoenvironmental reconstructions have limitations induced by multiple ecological assumptions. Knowing these assumptions remains essential to remember the limits of our interpretation, and using the Bioclimatic Analysis helps to partially overcome these limitations (Hernández Fernández, 2001; Hernández Fernández and Peláez-Campomanes, 2003, 2005; Royer et al., 2020).
The first limitation is the application of the principle of uniformitarianism, i.e. suggesting similar ecological tolerances between the past and present (Lyman, 2017; Pineda-Munoz et al., 2021). This principle is the first to be used when studying faunal remains. We do know, however, that many species have a high degree of behavioural and ecological plasticity or high level of endemism. Thus, some species may have changed and their ecological tolerances may have shifted. This is the reason why current paleoclimatic inference methods avoid considering only one species a unique indicator of one specific climate, and approaches use multiple taxa as a better means of reconstructing past environments. Using multiple species, as the Bioclimatic Analysis does, avoids then the overinterpretation related to one specific single taxon and allows then for a more nuanced understanding of climatic and environmental changes.
Second, the distribution pattern of modern biotic communities is the result of the combination of many factors (e.g. Jackson and Overpeck, 2000; Williams and Jackson, 2007; FAUNMAP, 1996), including geography (e.g. orography, refugia), biotic interactions (e.g. facilitation, competition) and history (e.g. climatic and/or geographic changes that cause population expansion or contraction, isolation, extirpation/extinction, and eventual speciation). This might lead to the development of past communities in which several species from contrasting environments lived, the so-called disharmonious faunas (Lundelius, 1989; Graham, 2005). The contrary effect is also found in modern assemblages, with some rodent species unexpectedly found in climatically very different environments, like the snow vole (Chionomys nivalis), which inhabits very high mountainous regions (e.g. the Alps and the Pyrenees) and lives equally in Mediterranean rocky inlets (Quéré and Le Louarn, 2011). In the same way, past refugia have played a fundamental role in the evolution of species, providing sites to survive extinction and to initiate colonization of new areas when environmental conditions became favourable (e.g. Tougard et al., 2008; Stewart et al., 2010; Royer et al., 2016; Baca et al., 2023a). The current period is a particular period when compared to all past periods, being the result of past history (Jackson and Overpeck, 2000; Graham, 2005; Williams and Jackson, 2007) plus anthropogenic activities. Anthropogenic activities strongly impact some rodents by offering opportunities for synanthropic species to develop, strongly modifying the realized niches of many other species (e.g. Pineda-Munoz et al., 2021), by constraining or facilitating their expansion. For instance, anthropogenic activities in Europe have favoured species inhabiting open environments through deforestation, such as the common vole, Microtus arvalis (e.g. Baca et al., 2023b), but have contracted significantly the range of the common hamster (Cricetus cricetus) in central Europe due to agricultural activities and landscape fragmentation (Surov et al., 2016). The Bioclimatic Analysis was established based on faunal associations from several typical distinct regions representing the different biomes found on Earth today, and the bioclimatic characterization of the rodent faunas remains largely unbiased, since considering the ecology of each species at biome-scale limits the species dependence effect to a particular environment. The combination of species interactions and their biomes also opens up the possibility of identifying special cases, related to taphonomical issues, possible particular ecotones that have occurred in the past or biomes that were not similar to present ones.
Using faunal associations implies large prediction errors in climatic reconstructions since, like plants, rodents are directly and indirectly (through the resources they use) dependent on multiple “bioclimatic” variables (e.g. temperature fluctuation throughout the diurnal cycle, insolation, seasonal distribution of temperature and precipitation, snow cover) that influence their behaviour and life cycle. For instance, lemmings inhabit environments where MTCO values can easily go down to −15 °C, but this variable has a reduced impact on these Arctic rodents because they live under the snow. The critical factors for them are then the quality of the snow cover and the protection it provides from air surface temperature. More importantly, in comparison with plants, rodents belong to an upper trophic level, being also highly dependent on vegetation, which serves as a food source and protection from climate and predators. Some species are very restricted to one type of vegetation, such as the wood lemming (Myopus schisticolor), which exclusively inhabits coniferous forest with a thick moss cover (Eskelinen, 2002; Calandra et al., 2015). Such links between vegetation and rodents imply that all abiotic (e.g. climate, CO2 concentration) and biotic (e.g. pest distribution, other consumers) parameters that affect vegetation subsequently impact rodents. Since the Bioclimatic Analysis is based only on correlations, and not causations, relating faunal components to climate variables, all these interrelations are indirectly taken into account.
3.2 Modern climate
3.2.1 Climate zone classification
The climatic classifications obtained using modern rodent associations almost perfectly capture the distribution of climatic regions of the Western Palearctic (Fig. 2). Only 15.9 % (n=25) of localities were not well predicted, which is in agreement with the apparent error (13 %) published by Royer et al. (2020). Nevertheless, most wrong predictions tend to be concentrated around ecotonal areas between different climate zones (Fig. 2), particularly in the transition regions between the temperate area (VI) and the boreal climate zone (VIII) in southern Scandinavia or between the desert (III) and the Mediterranean climate (IV) along the northern African coast and Middle East. Few of these localities show posterior probability values below 0.95, suggesting that the model has difficulties in distinguishing the mixed faunal associations between two climatic zones. A number of classification errors come from regions with ample climatic variations and highly diverse environments at local and regional scales, such as the Anatolian and Iranian plateaus. They include mountainous areas that are home to a very wide range of rodents with distinct ecological affinities, from deserts and semi-deserts (Rhombomys, Meriones) to steppes (Ellobius, Cricetulus) and woodlands (Apodemus), which leads to mixed rodent communities. The classification posterior probabilities do not systematically detect these mix associations, although some were found in northern Anatolia and in the Balkans. Similarly, mountainous regions may also be associated with classification errors (e.g. the Apennines), due to the mixture of lowland species (several species easily survive up to 2000 m) and the relatively few species typical of high mountain environments (Marmota). Finally, some errors may be associated with insularity, where faunal associations are the result of the process of Holocene island isolation and the introduction of species by humans. For instance, in Ireland the absence of many vole species and the presence only of the wood mouse and squirrel represent a clear case of out-of-equilibrium communities that act to bias climate classifications. Nonetheless, despite all these potential sources of errors (communities from ecotonal areas, highly heterogeneous areas or areas with peculiar biogeography history), the climate of 84 % of the modern rodent assemblages was correctly classified.
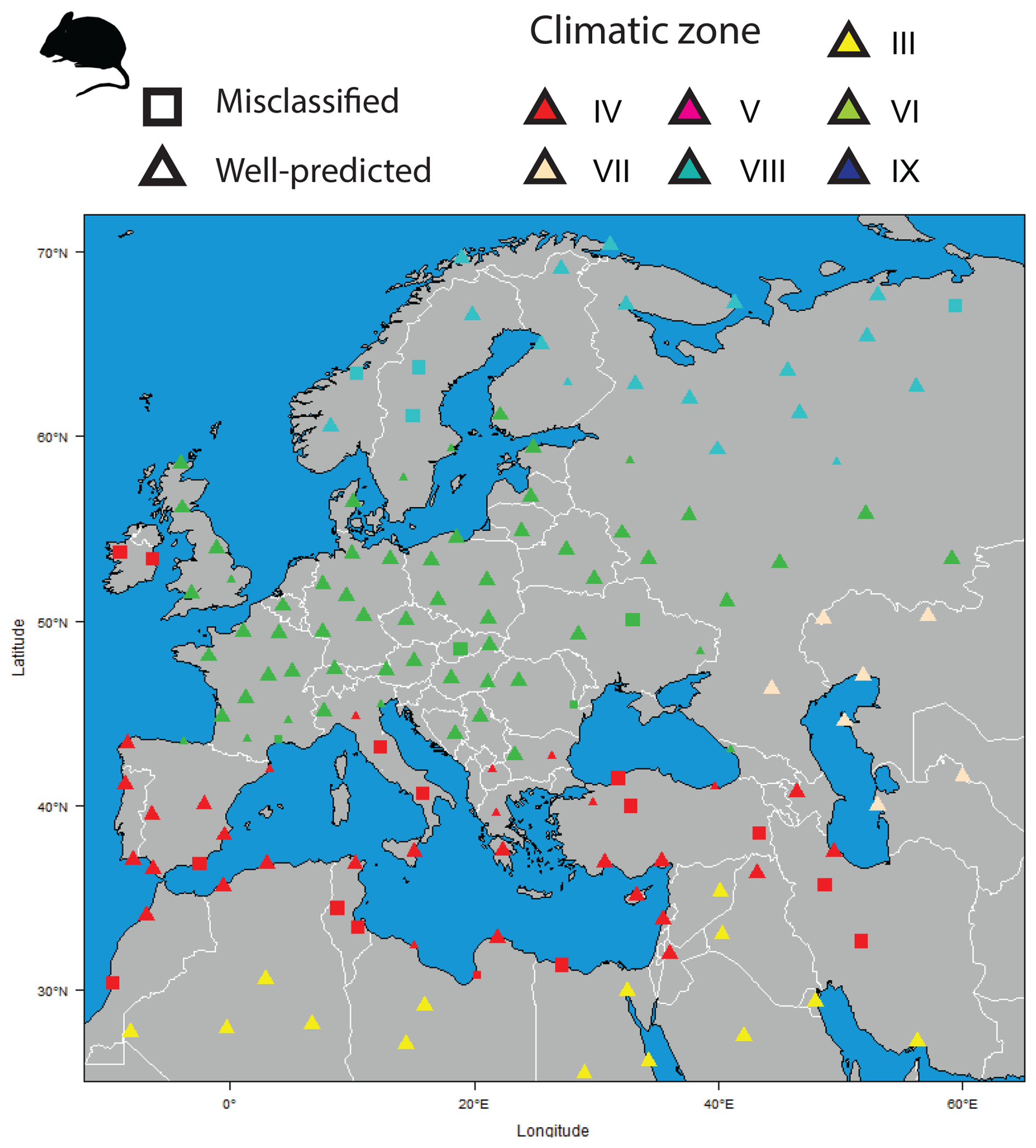
Figure 2Present-day climatic classifications obtained using modern rodent associations. Colours represent the climatic zone estimated by the Bioclimatic Model (see Table 1). Triangles represent well-classified localities, while squares depicts not well classified localities in comparison with their modern situation (see Table S1). These last symbols show two sizes, the larger one representing the well-established localities by Bioclimatic Model with posterior probability values higher than 0.95, while the small one presents localities with posterior probability values smaller than 0.95 obtained by the Bioclimatic Model.
3.2.2 Temperature interpolations
Our rodent-based reconstructions of modern MAT, MTWA and MTCO depict a geographical distribution that is consistent with the spatial pattern shown by GCMs, Beyer2020 and ERA5 (Fig. 3). The area-averaged temperature values estimated by rodent-based reconstructions reach 11.2 °C for the MAT, 23.6 °C for the MTWA and −0.4 °C for the MTCO (Table 2, Fig. 4).
Table 2Area-averaged MAT, MTWA and MTCO (°C) in rodent-based estimations, ERA5 and PMIP4 and Beyer2020 GCMs for different time periods covered by the data (present-day, LGM, Heinrich Stadial, Bølling, Allerød and Younger Dryas (YD)). The area average includes the 55 locations (see text and Table 2). Anomalies against the present-day conditions are also shown.
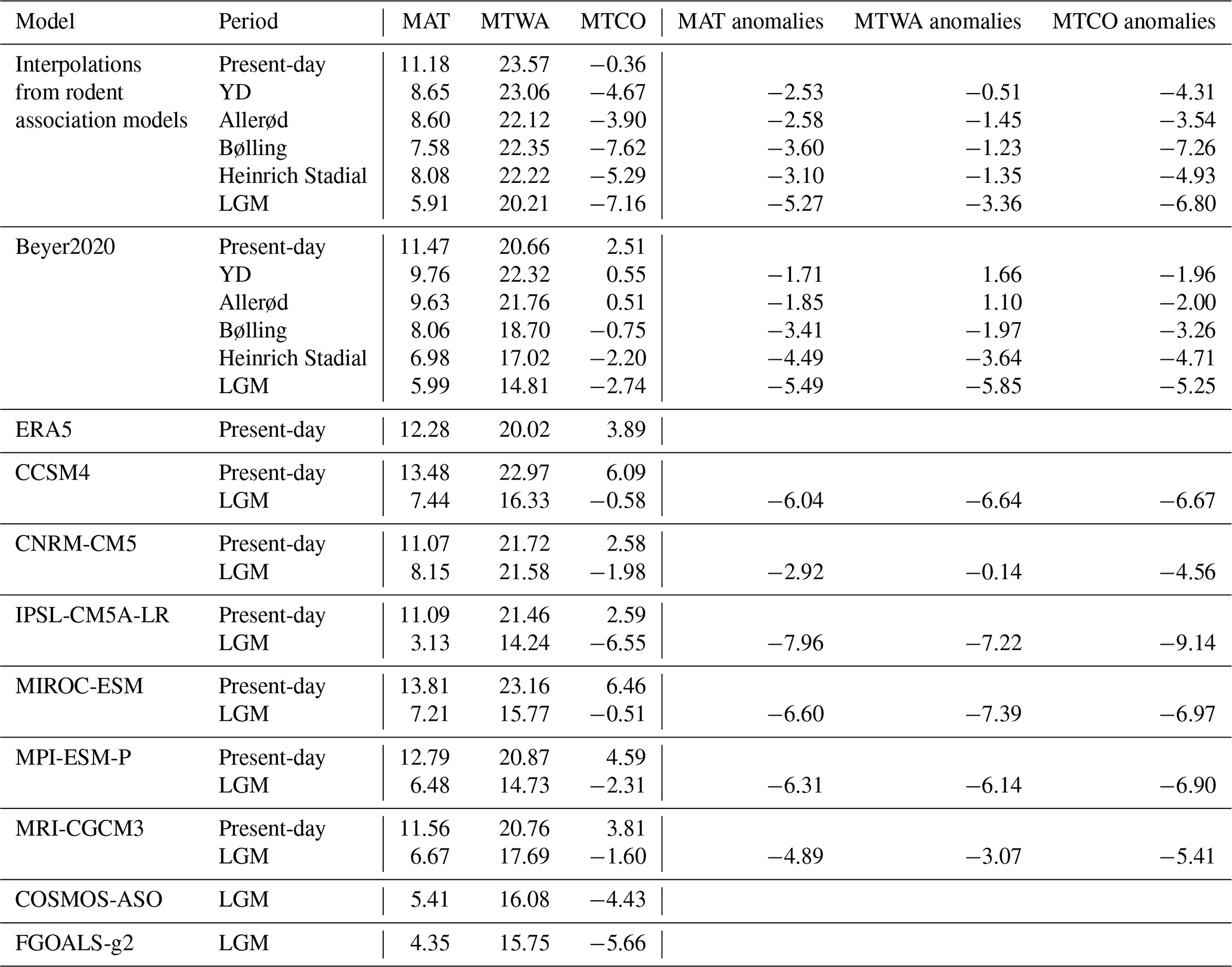
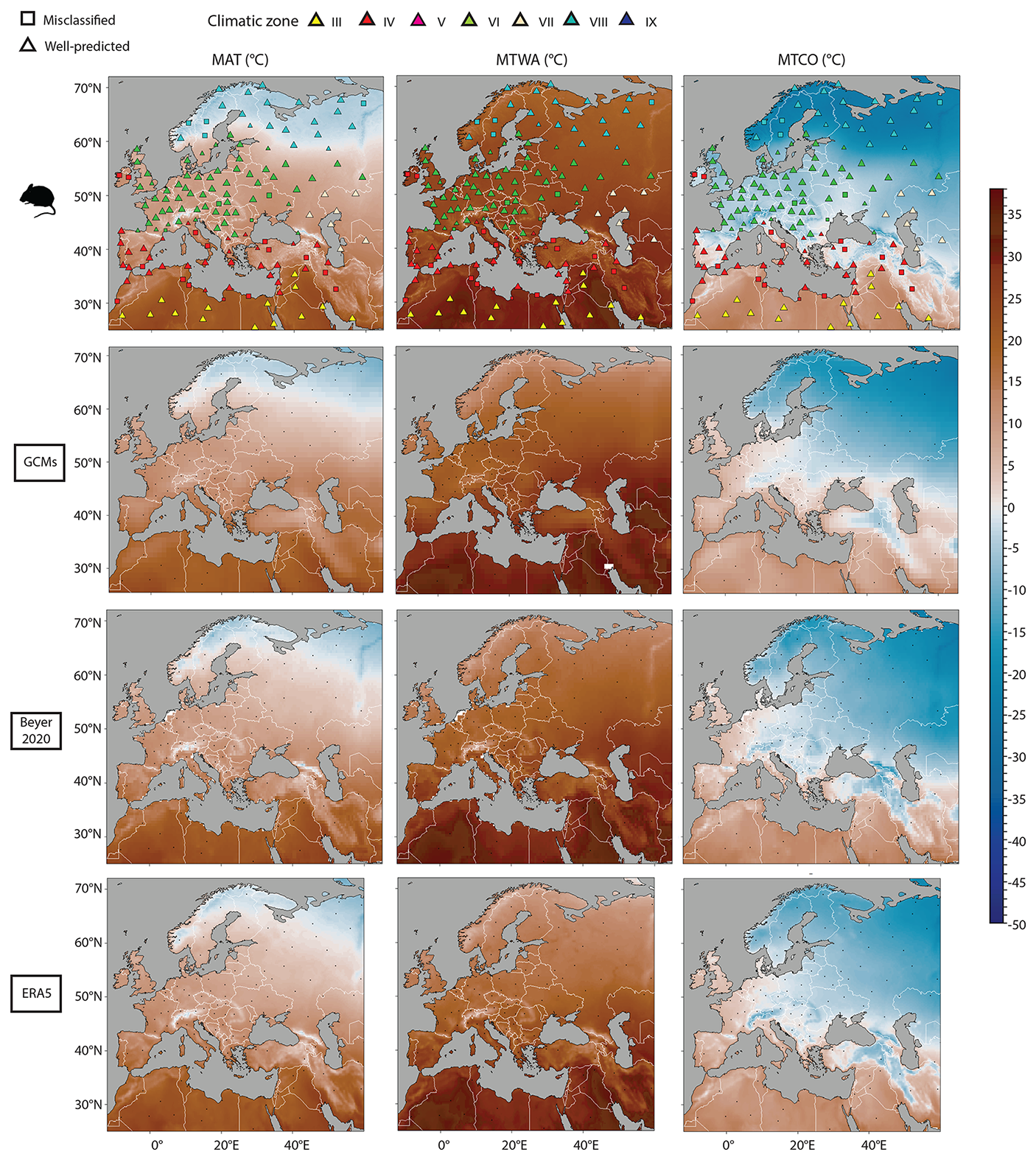
Figure 3Mean annual surface temperature (MAT), mean temperature of the warmest month (MTWA) and mean temperature of the coldest month (MTCO) estimated from 157 modern rodent associations spatially distributed all over the western part of Europe and the northern part of Africa (Fig. 1). For rodent predictions, colours represent the climatic zone estimated by the Bioclimatic Model (see Table 1). Triangles represent well-classified localities, while squares depicts not well classified localities in comparison with their modern situation (see Table S1). These last symbols show two sizes, the larger one representing the posterior probability values higher than 0.95, while the small one presents the posterior probability values obtained smaller than 0.95. For comparison, equivalent data are shown for ERA5 (restricted to 55° E), the PMIP4 GCM ensemble mean and the Beyer et al. (2020) GCM. Small black dots represent the localities with rodent associations.
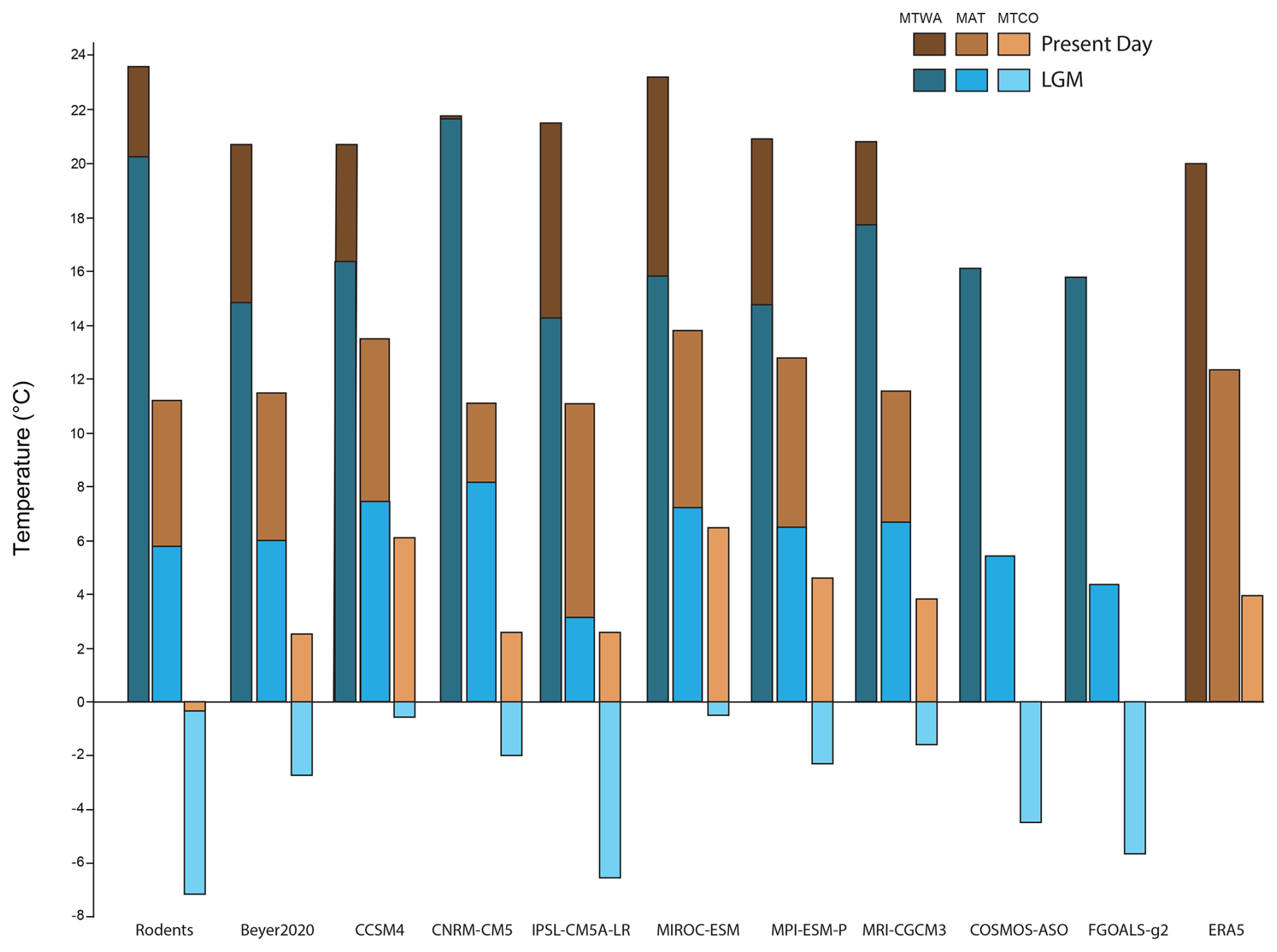
Figure 4Bar plot of area-averaged MAT, MTWA and MTCO for the present day and the LGM, as estimated from rodent associations, ERA5 and multiple GCMs. The area average is based on the 55 locations not affected by maximum ice-sheet extension (see text and Table 2).
The averaged MAT value is 1 °C lower than the ERA5 value and is very close to 4 of the 6 GCM values (Table 2, Fig. 4). At the site scale, despite the presence of few obvious outliers, the MAT rodent-based reconstruction values are within the GCM uncertainty range (Fig. 5a). The most sizable pattern for MAT is the abrupt decrease in temperature between climate zones VI and VIII, a pattern captured by our rodent-based reconstruction despite cold biases in climate zone VIII. The relationship between rodent estimations and ERA5 is strong (R2 = 0.92), comparable with the relationship between ERA5 and the GCMs (with R2 ranging between 0.87 and 0.97: Fig. 5b). MAT values obtained from the Bioclimatic Analysis of rodent assemblages show spatial discrepancies with ERA5 (Fig. 6). There is a tendency towards cold biases (∼ −3/−4 °C) all along the Atlantic coast latitudinal gradient, contrasting with positive biases (up to ∼ +3 °C) over Ukrainian and Russian steppes (Fig. 6). Similar bias patterns are observed when comparing ERA5 with GCMs and Beyer2020 estimations, but with weaker differences, as in the Iberian Peninsula or in Scandinavia (Fig. 6).
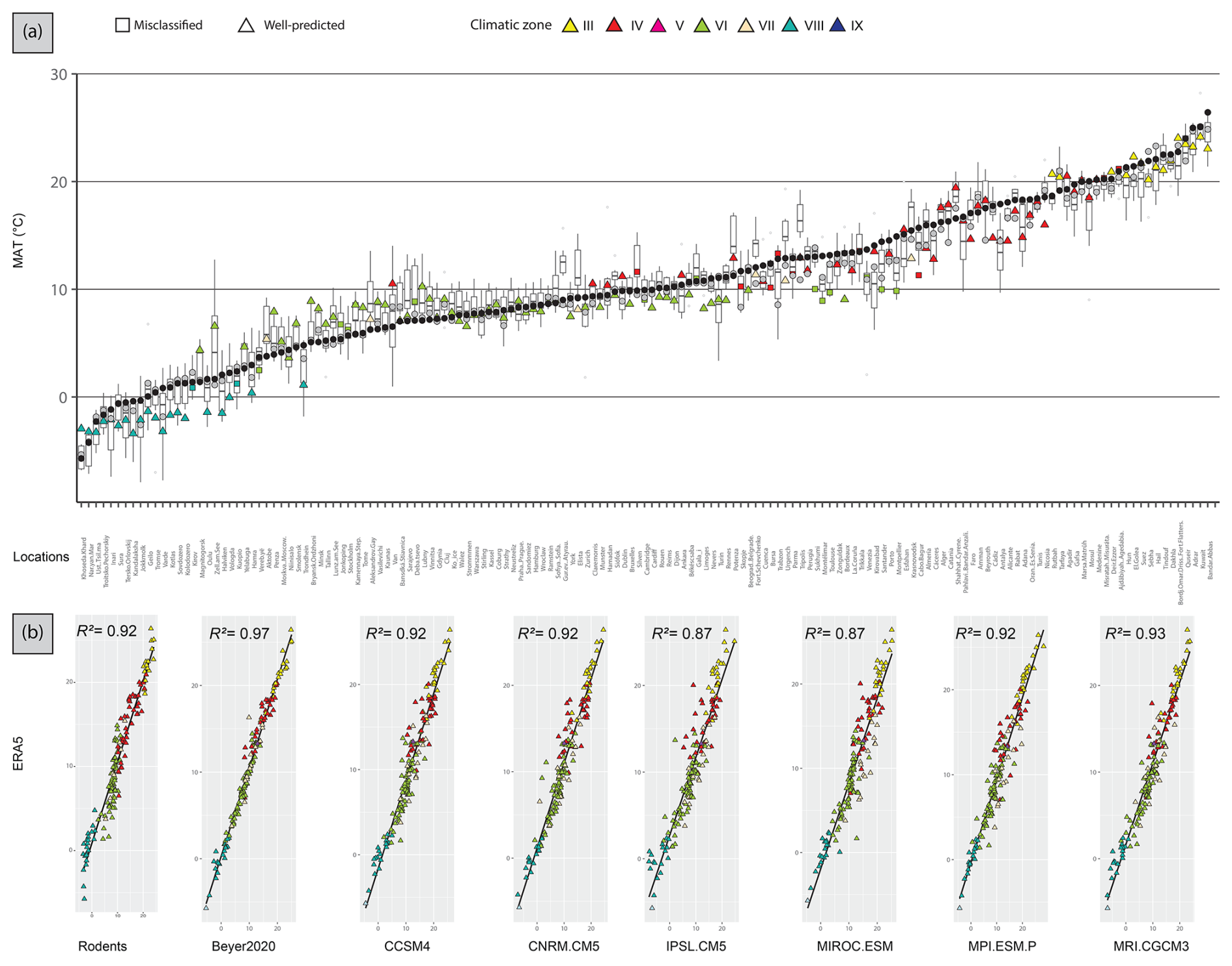
Figure 5(a) Present-day area-averaged MAT estimated from 157 rodent associations. Triangles represent localities whose modern climate zone was correctly classified, while squares are localities wrongly classified. Colours represent the climatic zone estimated by the Bioclimatic Model (see Table 1). MAT estimations from Beyer et al. (2020) (grey circles) and ERA5 values (black circles) are also shown. The boxplots represent MAT estimations from six different PMIP4 GCMs. Localities were ordered according to modern MAT values from ERA5 estimations. (b) Scatterplots and linear regressions between present-day MAT from ERA5 and the estimations from rodent assemblages or from the GCMs. All of them are statistically significant (p < 0.05) and R2 is shown.
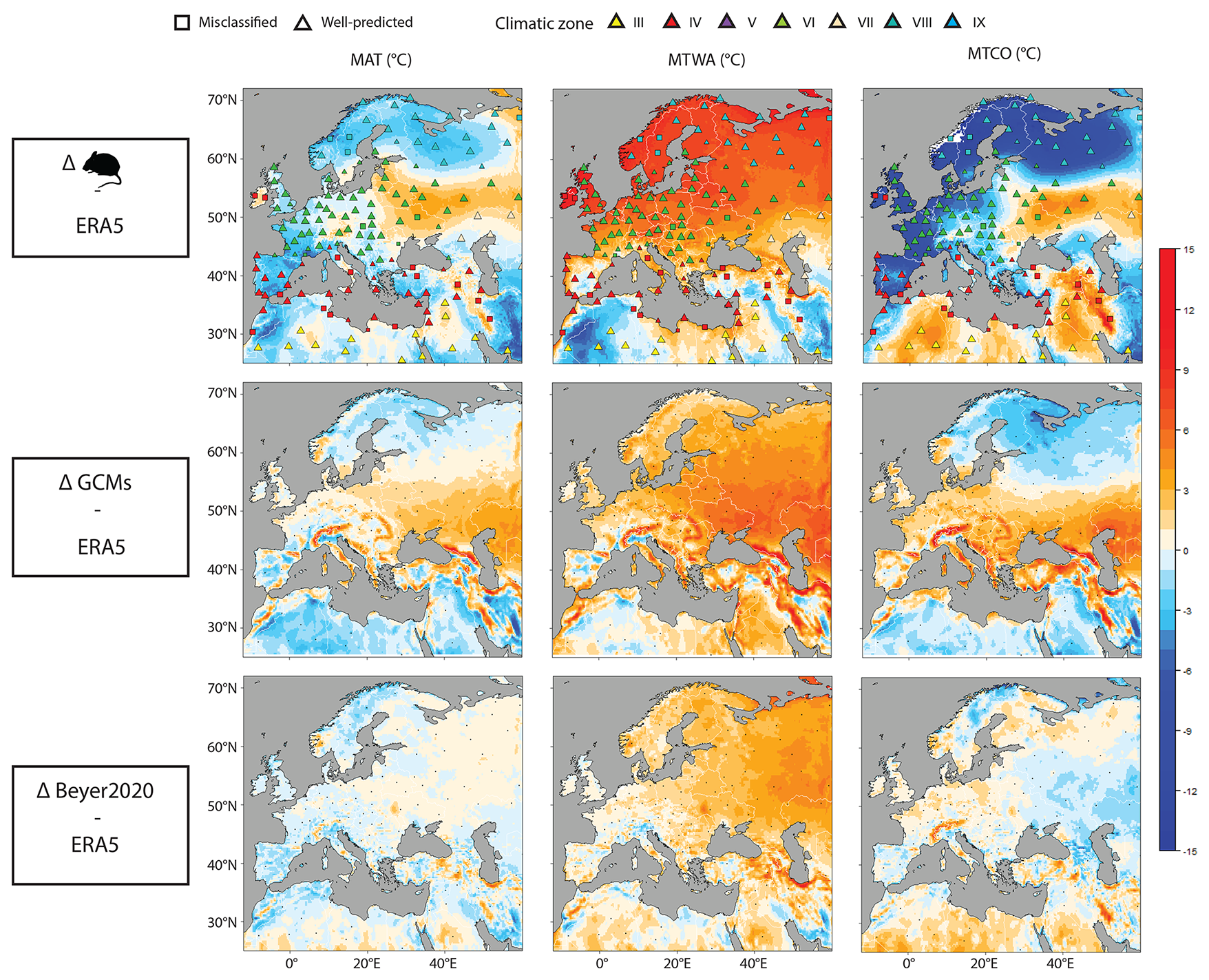
Figure 6Differences in MAT, MTWA and MTCO for present-day conditions between rodent-based, PMIP4 GCM ensemble mean and Beyer2020 GCM estimates and the ERA5 reanalysis. Warm colours indicate that the models estimate higher temperatures than ERA5, while cold colours indicate the opposite.
Compared to ERA5, MTWA values are overestimated by all models over Europe. The warm bias reaches almost 4 °C for rodent-based reconstructions, 2 to 3 °C for GCMs and less than 1 °C for Beyer2020 (Table 2, Fig. 4). At the site scale, although most rodent-based reconstruction values fall within the range of the GCM uncertainty, they are usually in the warmer range of their estimations (Fig. 7a). Rodent associations are generally used to infer warmer conditions than ERA5 and Beyer2020 for most of the climate zones, particularly for climate zones VI and VIII (Figs. 6 and 7a). Climate zones III and VII, on the other hand, show more accurate estimations (Fig. 7a). The regressions between ERA5 and the rodent estimates remain, however, still satisfactory (R2 = 0.84) as GCMs (R2 = 0.79–0.93), which indicates that the rodent-based model captures the spatial pattern of MTWA variation relatively well, although the slope and intercept go significantly away from 1 and 0, respectively (Fig. 7b). Discrepancies between rodent-based estimations and ERA5 show a similar pattern to GCMs biases, although somewhat more pronounced. It generally consists of higher temperature estimates, especially at latitudes above 50° N, and lower estimation under 40° N (Fig. 6).
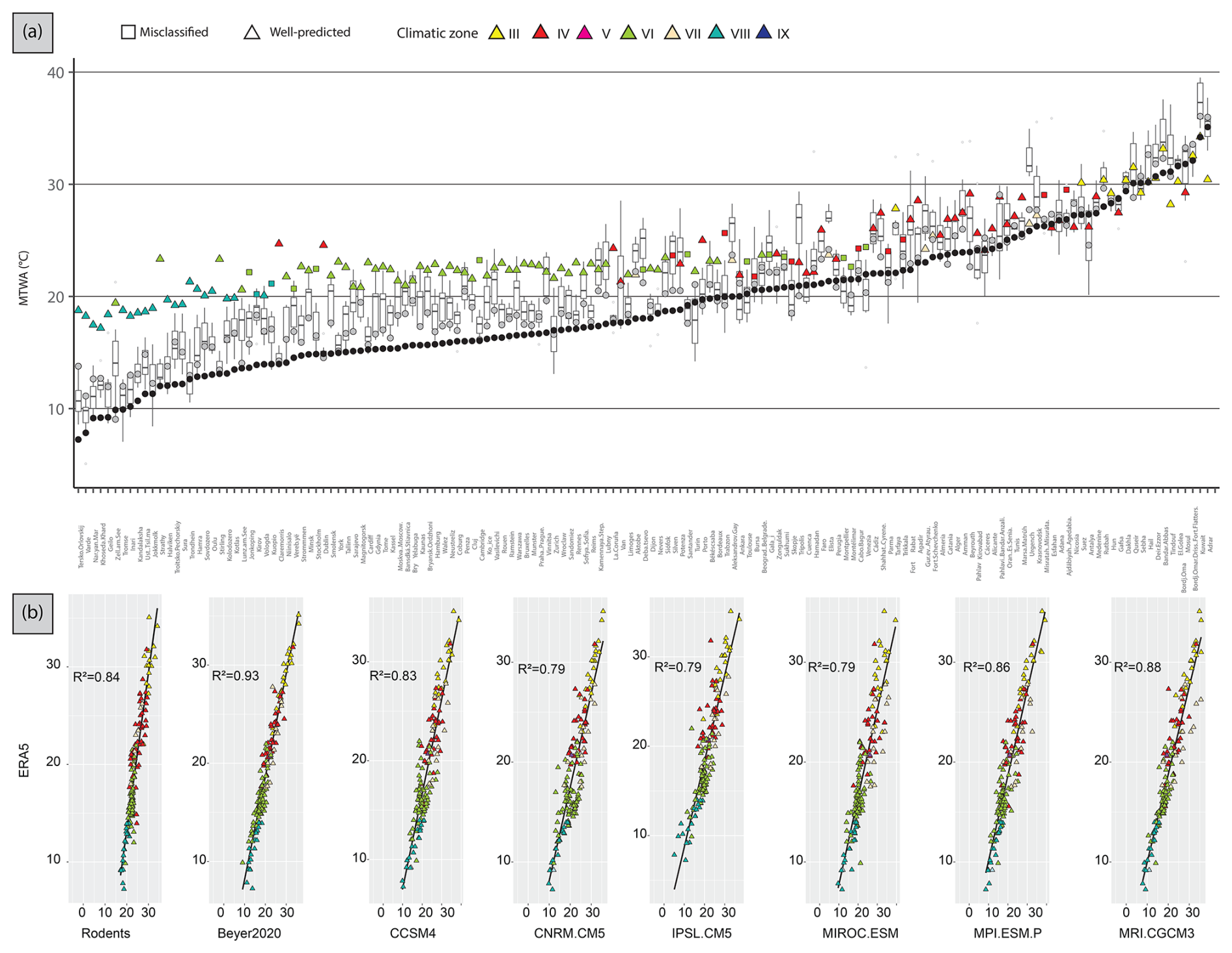
Figure 7(a) Present-day area-averaged MTWA estimated from 157 rodent associations. Triangles represent localities whose modern climate zone was correctly classified, while squares are localities wrongly classified. Colours represent the climatic zone estimated by the Bioclimatic Model (see Table 1). MTWA estimations from Beyer et al. (2020) (grey circles) and ERA5 values (black circles) are also shown. The boxplots represent MTWA estimations from six different PMIP4 GCMs. Localities were ordered according to modern MAT values from ERA5 estimations. (b) Scatterplots and linear regressions between present-day MTWA from ERA5 and the estimations from rodent assemblages or from the GCMs. All of them are statistically significant (p < 0.05) and R2 is shown.
The averaged MTCO value estimated from rodent associations is 4 °C lower than that of ERA5, and 2.8–6.7 °C lower than that of the GCMs (Table 2, Fig. 4). At the site scale, rodent-based estimations are also usually colder than the ERA5 and GCM values, in particular for sites located in climate zone VIII (Fig. 8a). The common variance between ERA5 and rodent-based estimations is weaker (R2 = 0.78) than with the GCMs (R2 = 0.85–0.96). This is mainly due to large discrepancies in sites located within temperate (VI) and boreal (VIII) climate zones (Fig. 8b). Spatially, southern locations are skewed towards warmer temperature inferences, without bias exceeding 5 °C. On the other hand, the Atlantic, Baltic and Arctic facades offer colder inferences (Fig. 6).
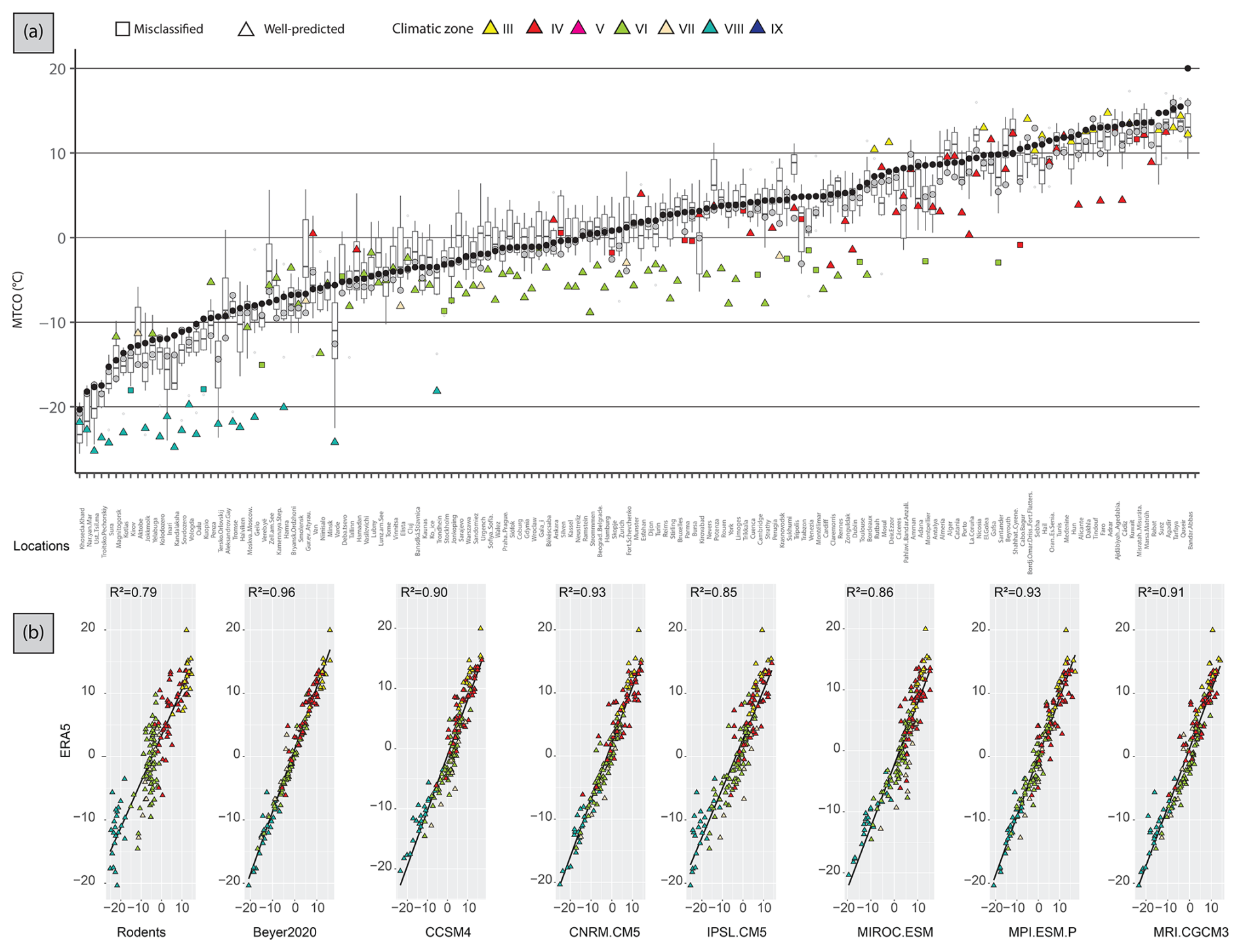
Figure 8(a) Present-day area-averaged MTCO estimated from 157 rodent associations. Triangles represent localities whose modern climate zone was correctly classified, while squares are localities wrongly classified. Colours represent the climatic zone estimated by the Bioclimatic Model (see Table 1). MTCO estimations from Beyer et al. (2020) (grey circles) and ERA5 values (black circles) are also shown. The boxplots represent MTCO estimations from six different PMIP4 GCMs. Localities were ordered according to modern MAT values from ERA5 estimations. (b) Scatterplots and linear regressions between present-day MTCO from ERA5 and the estimations from rodent assemblages or from the GCMs. All of them are statistically significant (p < 0.05) and R2 is shown.
To conclude, the evaluation of present-day spatial temperature patterns reveals a close relationship between small mammal communities and climate. Regressions between MAT, MTWA and MTCO and ERA5 show high R2 values, albeit slightly lower than the ones established by Royer et al. (2020). This is not surprising given the larger sample size used here, including localities from ecotonal areas among different climate zones. The rodent-based inferences accurately capture the “observed” (i.e. ERA5) spatial patterns of macroclimate, particularly in the case of MAT predictions. In the case of MTWA and MTCO, if global spatial patterns of macroclimate are well captured, estimations depict intensity biases in western and northern Europe (strong warm bias and cold bias, respectively), implying overestimated annual thermal ranges. These results indicate that the Bioclimatic Analysis (Hernández Fernández, 2001; Hernández Fernández and Peláez-Campomanes, 2003, 2005; Royer et al., 2020) has strong potentialities, not only to infer past climate conditions of isolated assemblages, but also to be integrated into paleoclimatic research based on interpolation at continental scale.
3.3 Past period reconstructions
3.3.1 The Last Glacial Maximum, a fistful of rodents
The climate zones inferred from LGM rodent associations (Fig. 9a) suggest that northern France and Germany experienced a boreal climate (VIII), in which tundra–steppe rodent species, such as collared lemmings (Dicrostonyx torquatus), narrow-skull voles (Stenocranius anglicus) or different ground squirrels (Spermophilus sp.), were particularly abundant (e.g. Hanquet et al., 2016; Markova et al., 2019). These rodents were part of the “mammoth steppe” or “steppe–tundra”, a uniquely high productive grassland ecosystem that supported a high biomass of large herbivores (e.g. horse, steppe bison, woolly rhinoceros and mammoth). These open landscape faunal associations are sometimes associated with woodland species, such as Clethrionomys sp., which suggests the presence of some discontinuous forest environments (e.g. Cyrek et al., 2000; Ponomarev et al., 2013; Nadachowski and Valde-Nowak, 2015). Only the rodent assemblage from Ireland supports a periglacial climate zone (IX), with communities entirely composed of lemmings (Woodman et al., 1997; Monaghan, 2017). Much more diverse rodent communities, including lemmings, narrow-skull voles, tundra voles, snow voles, marmots, ground squirrels, garden dormouse and wood mouse, indicate that the temperate climate zone (VI) covered southern France (Marquet, 1993; Royer et al., 2016; Crégut-Bonnoure et al., 2018) and northern and coastal areas of the Iberian (e.g. Cuenca-Bescós et al., 2009; Garcia-Ibaibarriaga et al., 2019) and Italian (e.g. Berto et al., 2017; Luzi et al., 2022a) peninsulas (Fig. 9a). As noted by García-Morato et al. (2024), in the Iberian Peninsula, the rodent communities from the southernmost areas reveal only minimal changes. In central and eastern Europe, the temperate climate zone (VI) seems to be associated with the interface around 50° N between the southern extension of the boreal climate zone (VIII), identified in the central Urals, and the arid-temperate climate zone (VII) found in its southern foothills. In the communities of the latter climate zone, dry-steppe species, such as the steppe lemming (Lagurus lagurus) or grey hamster (Nothocricetulus migratorius), became dominant (Danukalova and Yakovlev, 2006; Kosintsev and Bachura, 2013; Ponomarev et al., 2015). The Mediterranean climate zone (IV) was mostly restricted to southwestern Iberia as well as central and eastern Mediterranean shores, which served as a refuge for most Mediterranean species (e.g. Berto et al., 2019a; Ceregatti et al., 2023). The geographical distribution of biomes obtained from LGM rodent associations largely matches the one described by Markova et al. (2019) and Puzachenko and Markova (2023), which divided Europe into three latitudinal zonal assemblages during this time interval: periglacial tundra, periglacial tundra–forest–steppe and periglacial forest–steppe. Puzachenko and Markova (2023) suggest that differences between the western and eastern European faunal associations of the LGM could be related to more arid environments of the eastern regions and to the impact of Atlantic influences for the western regions. Finally, the LGM reconstructed by the rodent-based model is characterized by cold and dry climate conditions, with faunal communities constrained by the large presence of the Fenno-Scandinavian ice sheet. These faunal communities suggest the large presence of open steppic environment on a large part of northern Eurasia, as already suggested by pollen (e.g. Tarasov et al., 2000; Binney et al., 2017; Davis et al., 2022). This environment is most likely characterized by very fragmented forested cover and a scarcity of woodland refugia, which would be nonetheless associated with the occasional presence of some thermophilous species (Davis et al., 2022).
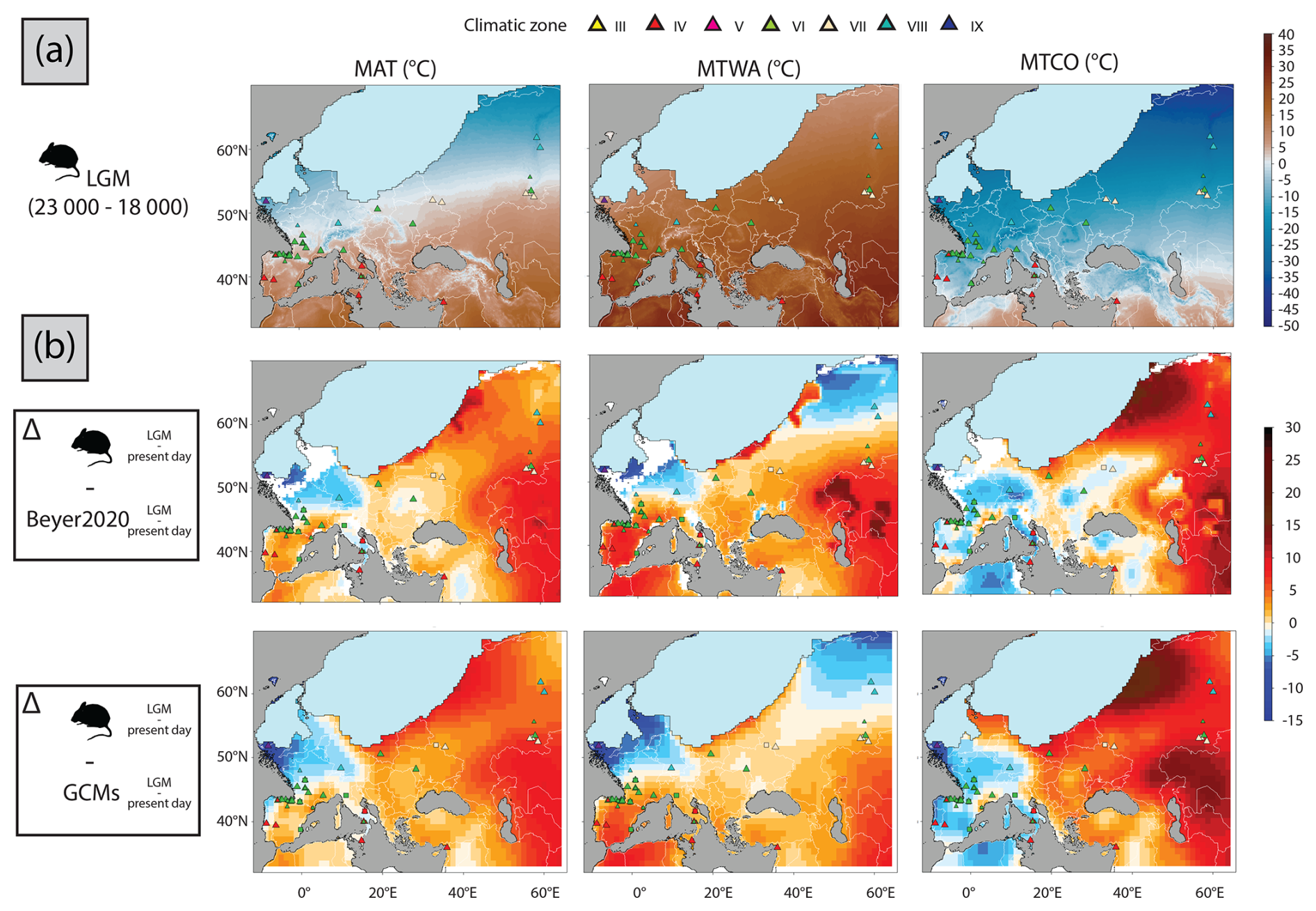
Figure 9(a) Rodent-based estimations of MAT, MTWA and MTCO for the LGM period. (b) Difference between the rodent anomaly values calculated between the LGM and the present-day periods (e.g. LGM minus the present day) and those calculated for Beyer2020 and from GCM models. Warm colours indicate that rodent-based estimations provide higher temperature changes than the PMIP4 or Beyer2020 GCM estimations, while cold colours indicate the opposite. Coloured triangles represent the climatic zone estimated by the fossil rodent associations using the Bioclimatic Model for the LGM period (Table 1). These symbols show two sizes, the larger one representing the posterior probability values higher than 0.95, while the small one presents the posterior probability values smaller than 0.95.
The temperatures estimated from the LGM rodent associations show a strong latitudinal gradient (Fig. 9a), with an average MAT anomaly of −5.3 °C compared with present-day temperature (Table 2). This is consistent with Beyer2020 (−5.5 °C) and within the range of uncertainty in PMIP4 GCMs (−3 to −8 °C). Rodent estimations from southwestern France range from 2 to 5 °C and are relatively similar to estimations from Beyer2020 (6 °C) and PMIP4 GCMs (3 to 11 °C). Rodent estimations reach around −3 °C at Brussels, while those from Beyer2020 are around 2.5 °C and PMIP4 GCM estimations range from −4 to 5 °C (Fig. S2). They are also 2–3 °C colder than other estimations based on faunal species (Puzachenko and Markova, 2023). Our rodent-based model suggests that the largest changes in MAT between the LGM and present-day conditions are observed in the Russian polar latitudes and around the British–Irish ice sheet (anomalies between −5 and −10 °C), while the cooling at lower latitudes was comparatively attenuated (anomalies between 0 and −5 °C) (Fig. 10), as also observed from pollen-based climate reconstruction (e.g. Bartlein et al., 2011; Davis et al., 2022). Spatial patterns from the rodent-based model thus exhibit a stronger latitudinal gradient during the LGM than today and differ from Beyer2020 and PMIP4 GCMs (Figs. 9b, 10 and Fig. S2), the latter proposing a less marked latitudinal gradient in MAT across the Western Palearctic. This leads to the rodent-based model suggesting that MAT anomalies between LGM and present-day conditions were higher than those of PMIP4 GCMs and Beyer2020 in the northwestern part of Europe (in blue in Fig. 9b) and smaller in the southwestern and eastern part of Europe (in red in Fig. 9b). Particularly notable differences in anomalies are observed in the eastern area for which Beyer2020 models suggest larger MAT changes than the rodent-based model does, as in the southern Urals, where rodent anomaly estimations are small; this is also probably emphasized due to the absence of rodent associations in the southeastern part of the study area for this time interval, which may produce spurious temperature inferences for lower southeastern areas (see red area in Fig. 10).
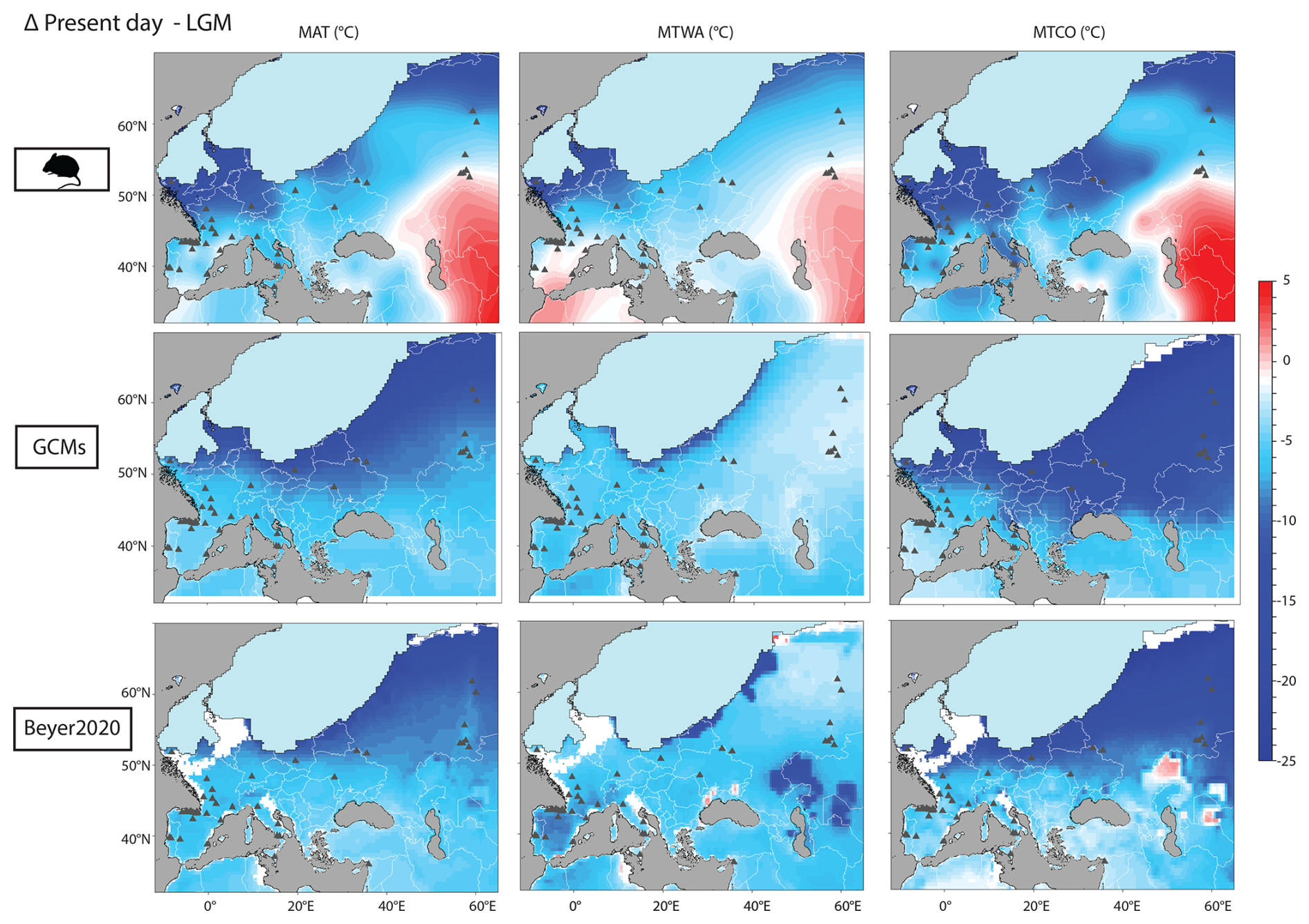
Figure 10MAT, MTWA and MTCO anomalies calculated between present-day and LGM conditions (LGM minus the present day) for the rodent-based, PMIP4 and Beyer2020 estimations. Cold colours indicate lower temperatures during the LGM compared to the present day, while warm colours signify the opposite.
Rodent-based changes in MTWA and MTCO between present-day and LGM conditions are similar to those obtained for MAT, although changes in MTCO are stronger than those in MTWA (Table 2, Figs. 10 and S2). On average the change in MTCO reaches almost −7 °C, while the MTWA anomalies only reach −3.4 °C (Table 2), with almost the lowest LGM to present-day changes for the southern latitudes. This is in agreement with Davis et al. (2022), who show that LGM climatic cooling was significantly greater in winter than in summer using updated LGM pollen estimations in Eurasia (Peyron et al., 1998; Jost et al., 2005; Wu et al., 2007). Our estimations are relatively close to those of Beyer2020 and PMIP4 GCMs (Table 2). However, spatial patterns differ significantly between the rodent estimations and the other models. Rodent-based estimations of MAT, MTWA and MTCO are colder in the northwest region and warmer in the eastern region compared to Beyer2020 and PMIP4 GCMs (Fig. 9b).
Figure 10 summarizes temperature changes between present-day and LGM conditions across all models and indicators. A pronounced meridional gradient is evident in all models and for all indicators, with stronger cooling in the north than in the south. In addition to this meridional gradient, the rodent model reveals a geographical gradient, showing LGM temperatures that are much colder in the northwest and slightly warmer in the southeast compared to present-day conditions. This pattern is consistent across MAT, MTWA, and MTCO but does not appear in Beyer2020 or the GCMs. It is linked to the presence of an Arctic rodent association in the northwest, possibly indicating lower heat input associated with a weak Atlantic Meridional Overturning Circulation (AMOC). The lack of this zonal gradient in GCMs may be due to an overly strong AMOC, despite improvements from PMIP3 to PMIP4 (Kageyama et al., 2021a).
As noted by Davis et al. (2022), although the traditional view of the LGM has been defined for many years, many questions still remain to be explored when more precise quantification is researched. The LGM is depicted as a very different period compared to today, with distinct climate, vegetation and landscapes, together with a different orographic context including large ice sheets, which have fully impacted faunal communities. Typical cold rodent species have been recovered throughout mid-latitudes and high latitudes and were mainly limited by natural barriers like mountains from reaching southern latitudes. They form ecotone rodent communities, mixing species from polar, boreal and temperate environments, and are far from reflecting typical full periglacial communities, except for the ones located at the closest range of the ice sheet. The quantification obtained with the rodent-based model supports the global view of a colder LGM world, but with MAT anomalies no larger than −5.5 °C compared to the present-day conditions, and underpins the finding that changes in MTCO were stronger than in MTWA. Nonetheless rodent associations produce spatial temperature patterns different than the ones estimated by Beyer2020, suggesting notably greater temperature changes in the northwestern part of Europe, while the temperature anomalies in the southwestern and the eastern part of Europe were weaker than those of Beyer2020 and PMIP4 GCMs.
3.3.2 Once upon a time in the Late Glacial
Compared to the LGM picture, the Heinrich Stadial experienced sizable changes in species distribution throughout the Western Palearctic (e.g. Royer, 2016; Royer et al., 2016; Markova et al., 2019). However, climate zone distribution remains globally similar between these two periods (compare Fig. 11 with Fig. 9a). Northern Europe yielded more sites with Arctic rodent associations, like in Belgium, Poland and the northern Ural Mountains (Teterina, 2009; Stewart and Parfitt, 2011; Ponomarev et al., 2012; López-García et al., 2024), forming a composite mix of boreal (VIII) and polar (IX) climatic zones. The transition between climatic zones VI and VIII appears south of 50° N in the western part of Europe and around 55° N in the eastern part, while central Europe remains dominated by an arid-temperate climate zone (VII), as shown in Fig. 11. The Iberian and Italian peninsulas show a mix of both Mediterranean (IV) and temperate (VI) climate zones. With the warming of the Bølling and the Allerød, large changes occurred at the species level (Kalthoff, 1998; Jeannet, 2009; Royer et al., 2016, 2021; Rofes et al., 2020; Wong et al., 2017). Changes in the climate zone distribution occur slightly with the boreal climate zone (VIII) in western Europe retreating towards northern latitudes as the ice sheet melts. The temperate climate zone (VI) slightly develops to the north up to 50° N, while central and eastern Europe seems to present a pattern relatively similar to previous intervals with the presence of an arid-temperate climate zone (Fig. 11). Yet, to date, there is still a general lack of information on the southeastern part of the study area for this time interval, making it difficult to understand how exactly the arid-temperate climate zone (VII) has changed. The Iberian and Italian peninsulas still deliver rodent associations of both Mediterranean (IV) and temperate (VI) climate zones, although the latter are becoming rare. There are few fossil sites to cover the Younger Dryas, and most of them are in western Europe. They do not reveal remarkable changes in climate zone distribution, despite the cooling of this event. The British Isles still experienced boreal (VIII) and polar (IX) climate zones, attested by the presence of typical boreal and polar lemming (Price, 2003; Arbez et al., 2021), while a boreal climate zone was established in the northern part of the Urals (Fig. 11).
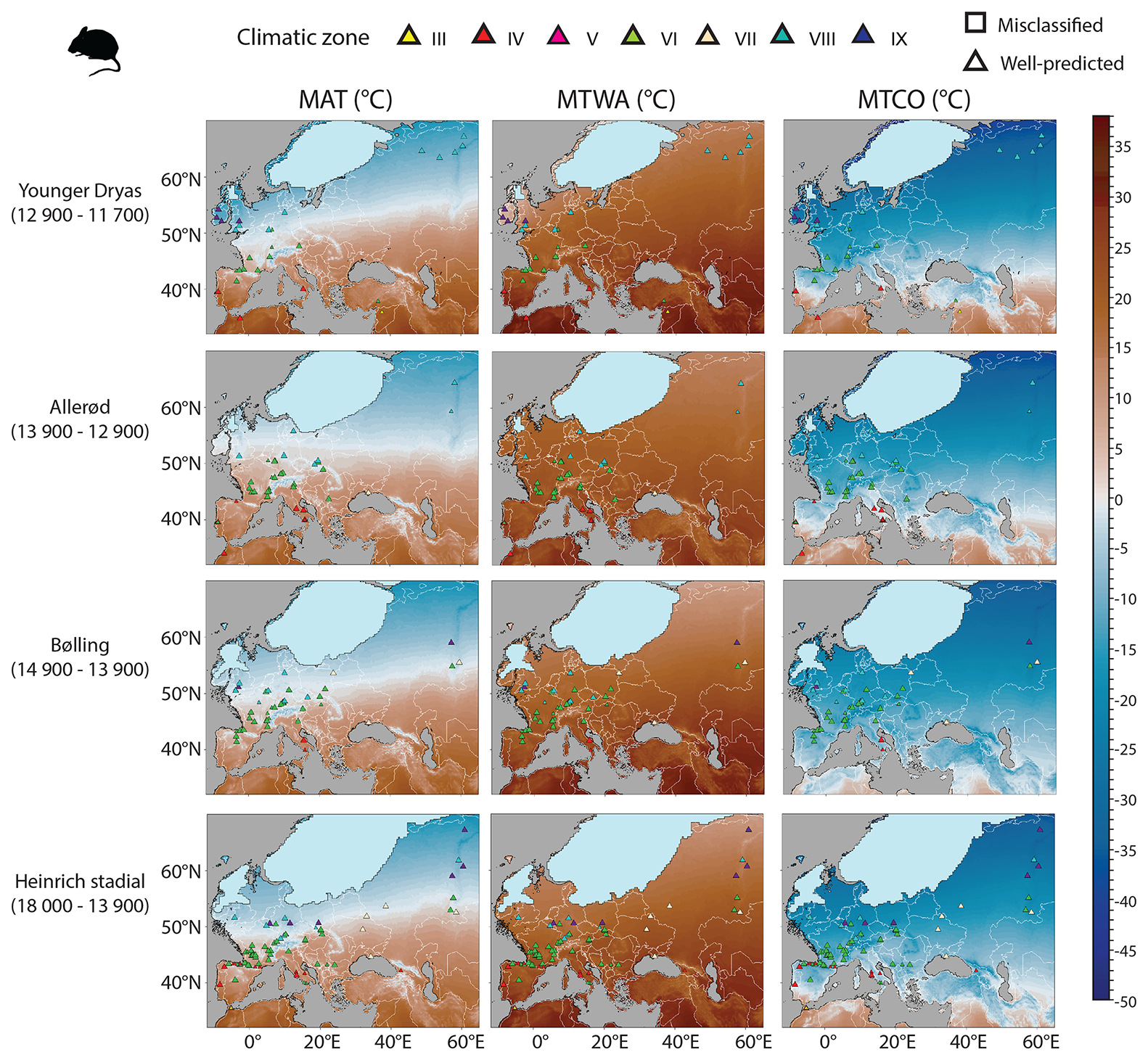
Figure 11Rodent-based estimations in MAT, MTWA and MTCO for the Heinrich Stadial, Bølling, Allerød and Younger Dryas periods. Coloured triangles represent the climatic zone estimated by the fossil rodent associations using the Bioclimatic Model (Table 1). These symbols show two sizes, the larger one representing the posterior probability values higher than 0.95, while the small one presents the posterior probability values smaller than 0.95.
Average climate estimations obtained from rodent associations present a gradual temperature warming between the LGM and the present day (Table 2). MAT of different Late Glacial events remains however within a relatively narrow range of values (7.6 to 8.7 °C), with the MAT value at 8.1 °C since the Heinrich. MTWA values vary little (between 22.1 and 23.1 °C), while MTCO values range from −7.6 to −3.9 °C. These estimations obtained with the rodent-based model are relatively close to those of Beyer2020, in particular for MAT (Table 2). For MTWA and MTCO, Beyer2020 estimations suggest larger temperature changes across the events, while rodent-based reconstructions suggest only small changes in MTWA and still larger MTCO anomalies for Bølling–Allerød–Younger Dryas periods (Table 2).
These spatial temperature reconstructions of Late Glacial events differ from Beyer2020 in the same way (Fig. 12). Except the southeastern part for which no rodent association is available, the main disagreements come from a different latitudinal gradient of changes in the western part of Europe and differences in northeastern part of Europe. For the northwestern part of Europe, the rodent-based model estimates that MAT, MTWA and MTCO anomalies between Late Glacial events and present-day conditions were systematically higher than Beyer2020 (in blue in Fig. 12), thus suggesting larger temperature anomalies throughout all the Late Glacial. For instance, for the Bølling and the Allerød, MAT anomalies from the rodent-based model are estimated at around −12.5/−8 °C in England, compared with −5/−3.5 °C for the Beyer2020 model. By contrast, the rodent-based model yields smaller temperature anomalies for the southwestern part of Europe, as well as for the eastern part of Europe during the Heinrich Stadial and Bølling. For instance, MAT anomalies of the Heinrich Stadial and Bølling in southwestern France are estimated at around −3/−3.5 °C for the rodent-based model and are thus relatively close to the values of Beyer2020, estimated at −4.5/−5 °C. For the northeastern part of Europe, Beyer2020 estimated greater anomalies for MAT and MTCO between Late Glacial events and the present day (in red in Fig. 12) than the rodent-based model did. For the region of Moscow, MAT anomalies of Beyer2020 range from −13.5 to −11 °C for the Heinrich Stadial and Bølling, while the rodent-based model estimated MAT anomalies between −9 and −10 °C.
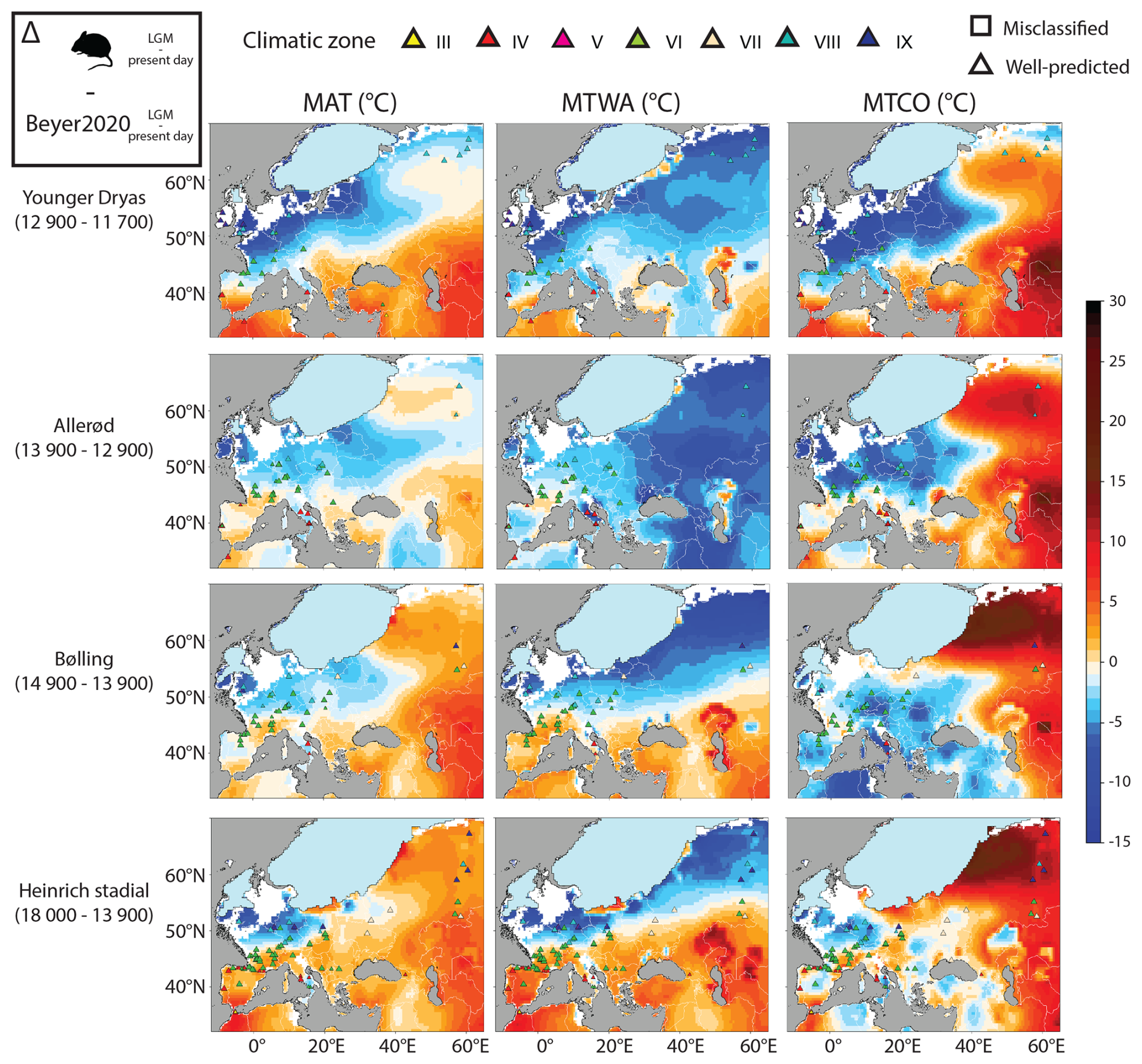
Figure 12Difference between the rodent anomaly values calculated between the past (Heinrich Stadial, Bølling, Allerød and Younger Dryas) and the present-day periods (e.g. LGM minus the present day) and those calculated for Beyer2020 models. Warm colours indicate that rodent-based estimations provide higher temperature changes than Beyer2020 estimations, while cold colours indicate the opposite. Coloured triangles represent the climatic zone estimated by the fossil rodent associations using the Bioclimatic Model for the past period (Table 1). These symbols show two sizes, the larger one representing the posterior probability values higher than 0.95, while the small one presents the posterior probability values smaller than 0.95.
As a consequence, temperatures obtained from rodent-based reconstructions are far from changing uniformly across Europe between the Late Glacial events (Fig. 13). The rodent-based model shows larger spatial variations than Beyer2020 temperature changes. Indeed the Beyer2020 model shows more spatially uniform changes for the MAT across Europe between the Late Glacial events, with the main changes located in areas close to the ice sheet, while some longitudinal variations have been observed for MTWA and MTCO estimations (Fig. S3). Conversely, the rodent-based model suggests greater latitudinal and longitudinal variation in changes. After the LGM, the increase in MAT rodent-based estimations during Heinrich Stadial is larger along the Atlantic seaboard than in central Europe. Southwestern France and the north of Spain experienced a warming of between 1.5 and 3 °C, while temperature changes are lower than 1 °C in Belgium, Germany and Poland. The northeastern part shows an increase of 3 and 6 °C for MAT and MTCO, respectively. Beyer2020 suggests a general rise of around 2 °C for MAT, with an increase no larger than 1.5 °C in southwestern France and Spain and between 1.2 and 5 °C in Belgium, Germany and Poland. During the Bølling, MAT and MTCO values fall within those of the rodent-based model for the middle and the lower latitudes, while they rise in the northwest, as in Belgium and Germany, where a MAT increase of 2 to 3 °C is found. The Allerød presents sizable changes, with a MAT warming between 1 and 3 °C in all of western Europe. The southern latitudes are mainly impacted by an increase in MTCO, with for instance an increase of 3.5 °C for the north of Spain and of 2 °C for southwestern France. The northern latitudes show an increase in MTWA, reaching up to 2.5 °C as in Brussels. The Younger Dryas event brings back well-marked glacial conditions before the beginning of the current interglacial, particularly along the Atlantic coast and the British Isles. The latter experienced a MAT decrease exceeding 5 °C (Fig. 13), while Beyer2020 shows no sizeable changes in this areas, except in Scotland, with a MAT increase of around 2.5 °C (Fig. S3).
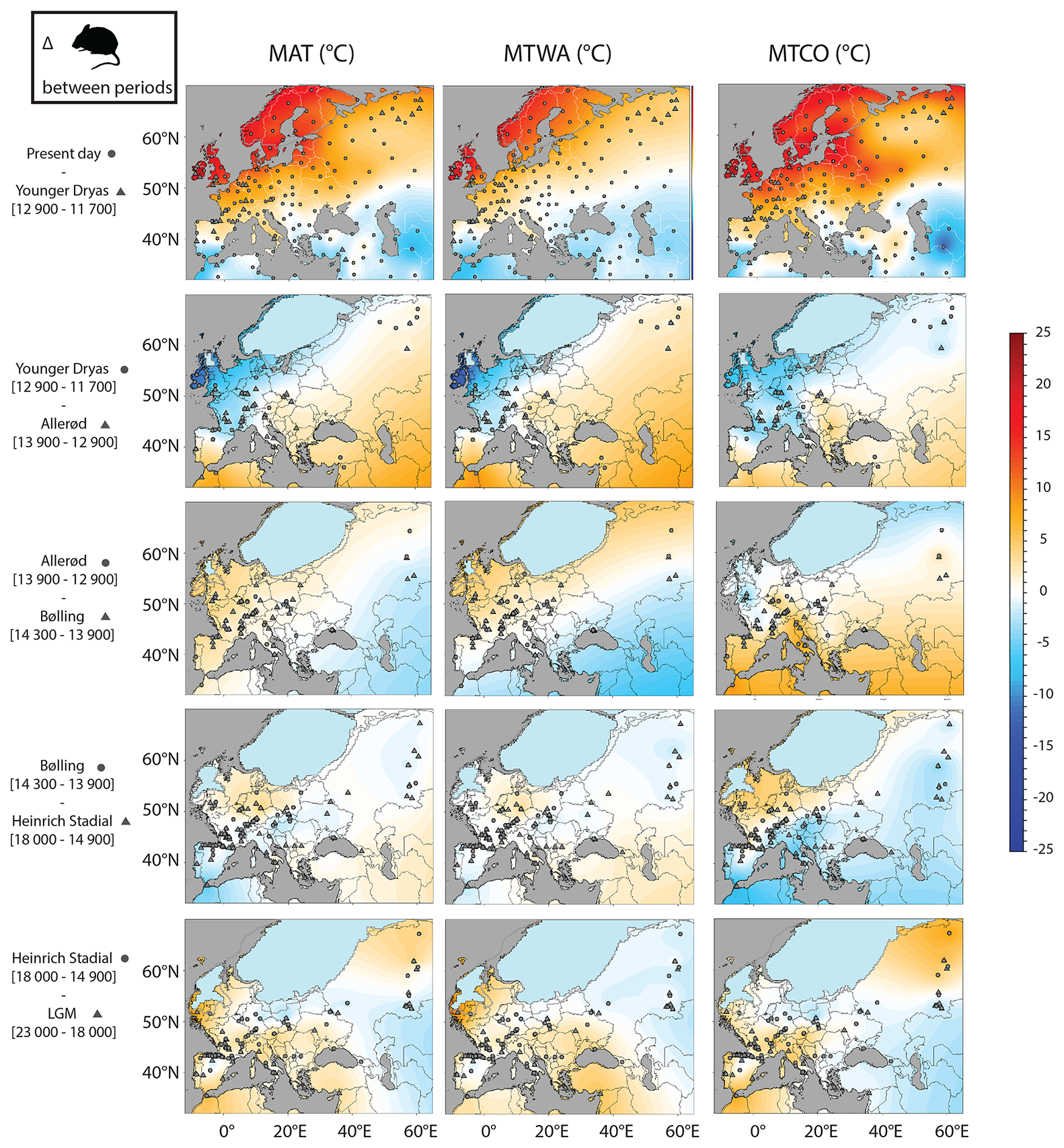
Figure 13Differences in MAT, MTWA and MTCO between each successive time period in rodent-based estimations. Triangles and circles represent the positions of fossil localities used in each pair of time intervals. The ice sheet of the most recent period is represented in blue, while the limits of the older one appears in grey.
The Late Glacial period experienced significant climatic shifts, marked by rapid climate oscillations, which led to a dynamic re-shaping of living spaces, with the progressive retreat of ice sheets freeing up new areas in the northern regions while rising sea levels simultaneously submerged substantial portions of coastal regions. These changes have restrained other areas, isolated some islands and induced significant changes in plant communities. All these biotic and abiotic changes have led to considerable changes in faunal associations throughout Europe, triggering range shifts, local extirpations, extinctions, population replacement, and isolation in refugia or recolonization processes (e.g. Stewart et al., 2010; Brace et al., 2016; Baca et al., 2017; Puzachenko and Markova, 2019; Magyari et al., 2022). At the community scale, the changes remain slow, as shown by the progressive rise of the temperate climate zone (VI) throughout the Late Glacial (Fig. 11) and in the early Holocene for the northern latitudes. In southwestern France, the changes in small mammal associations are gradual, taking all of the Late Glacial events to switch progressively from the typical glacial community to the typical interglacial community (Royer et al., 2016), including also the presence of certain species only at certain events (Royer, 2016; Royer et al., 2018). These Late Glacial periods therefore produce rodent associations mixing typical taxa of northern environments, with new southern and eastern taxa (Royer et al., 2016), which gradually evolve in latitude in line with the events (e.g. Cordy, 1991; Wong et al., 2020; López-García et al., 2024). These faunal changes reflect temperature changes that vary in both time and space, producing a spatial pattern that differs from that of Beyer2020. Rodent results suggest that the northwestern part of Europe experiences larger temperature changes, while temperature anomalies in the southwestern and the eastern part of Europe were smaller than those estimated by Beyer2020.
3.3.3 How to interpret the differences between GCMs and rodent-based inferences
As described previously, the rodent-based model provides patterns similar to those of GCMs for both present-day and past conditions, with for instance a basic picture of the LGM as a cold world with MAT anomalies around −5.5 °C in the Western Palearctic. Yet, there are notable divergences among the approaches. These divergences are generally included in the range of GCM uncertainties and hide sizable regional differences, like between northwestern and northeastern Europe (Figs. 9b and 10). Compared to GCMs, rodent associations of LGM and Late Glacial periods generate colder temperature values in the western part of Europe and warmer values in eastern Europe (Fig. 9b). This leads to a stronger west–east temperature gradient in the rodent-based model against both PMIP4 GCMs and Beyer2020, especially for the coldest months.
The physical interpretation of these distinct temperatures observed for past period is questionable. On the one hand, working with faunal remains requires taking into account a few taphonomical, analytical and ecological assumptions detailed in Sect. 3.1. The discrepancies between our rodent-based model and PMIP4 GCMs and Beyer2020 in higher latitudes (Fig. 10) could be explained by a limitation in the Bioclimatic Analysis. Therein, MAT of the coldest areas used to generate the new version of the Bioclimatic Analysis reaches around −15 °C (Royer et al., 2020) in areas with typical Arctic species such as lemmings or tundra shrews. It is then very difficult to properly extrapolate MAT values below −15 °C, while some GCM estimations easily reach up to −30 °C and even colder temperatures in eastern regions, bringing into question how faunal species identified through the fossil record could have survived in such conditions.
On the other hand, GCMs have their own uncertainties, biases and sensitivity to changes in the radiative forcing and boundary conditions. Each GCM uses different sets of parameterizations to approximate sub-grid processes that cannot be resolved at coarse spatial and temporal resolutions, which strongly influence climate simulations (Taylor et al., 2012; Varela et al., 2015), resulting in a large range of uncertainties across GCMs. GCMs often diverge from actual paleontological data (Jost et al., 2005; Allen et al., 2008; Braconnot et al., 2012; Latombe et al., 2018). The Beyer2020 model appears to propose the best estimation for the modern context (Figs. 4, 5, 7 and 8) due to bias correction. Interestingly, the authors evaluated their method for the past period at the global scale, comparing it only to a mid-Holocene and LGM pollen database (Bartlein et al., 2011). The question may arise regarding the validation of this model during unstable glacial periods when relying solely on a single proxy. This has been recently criticized because some of the pollen sequence was not necessarily properly attributed to the LGM (Davis et al., 2022). Furthermore, LGM is a peculiar period of the last glaciation, with a specific abiotic context. No validation has been tested for the other glacial periods, which present their own abiotic particularities.
On the basis of all these assumptions, we can finally conclude that these divergences between GCMs and rodent-based estimations could result from multiple factors that need further investigation. Beyond GCM limitations, our interpolations are “a point of view” of rodent associations, which could contribute to explaining the disagreements with GCMs. Species association may exist under environmental conditions that do not exist today but did exist in the past (Jackson and Overpeck, 2000; Williams and Jackson, 2007; Lyman, 2017). The relationships between the species and their environment are in constant evolution, and the current observations reflect only a realized niche, potentially underestimating the plasticity and adaptive capacity of species in response to new abiotic and biotic constraints. Rodent-based estimations tend to show a slightly different zonal temperature gradient from the GCMs, with colder values in western Europe and warmer values in eastern Europe (Figs. 9, 10 and 12). This gradient may be slightly amplified by the lack of spatial data in some regions. In the present case, some errors can be explained by the absence of close fossil sites (e.g. near the Scandinavian ice sheet) or their limited number (as in the British Isles). Our rodent estimations give MAT values in eastern Europe that are warmer than those provided by GCMs for all the time intervals of the Last Glacial period. These differences could also be related to differences in faunal communities and their paleoecology. The glacial world of the LGM and Late Glacial could not be simply compared to the current interglacial biomes. The very nature of these particular environments is not comparable due to their unique abiotic parameters like diminished solar radiation and the CO2 atmospheric concentrations and biotic factors such as the important presence of a thriving mammalian megafauna. This megafauna actively shaped the European vegetation structure (Malhi et al., 2016; Pearce et al., 2023; González-Varo, 2025), potentially driving the diversification of distinctive rodent communities.
Rodent associations are a regularly used proxy to reconstruct local past climate and environment contexts, as they are often found in archaeological and paleontological excavations and are highly sensitive to environment changes. Here, we focused our study on large spatial scales, over the Western Palearctic, to access continental-scale paleoclimate reconstructions based on a total of 279 archaeological and paleontological levels. Although there are entire regions with few or no data, our dataset encompasses a wide range of latitudinal gradients, providing distinct climatic contexts, which give us the opportunity to address the spatial background for changes in past climate conditions across the region.
The rodent-based climate reconstructions allow for exploring spatial changes between present-day conditions and different time periods ranging from the Last Glacial Maximum (LGM) to the end of the Late Glacial (i.e. 23 000 to 11 700 years before present). Our main findings highlight the following about rodent-based reconstructions:
-
They are efficient for examining large spatiotemporal variations in past climate, enabling detailed investigations of temperature changes at continental scale, since they accurately capture the modern distribution of climatic zones in the Western Palearctic, as well as (ERA5) spatial patterns of macroclimate, notably for MAT.
-
They exhibit a stronger latitudinal gradient during the LGM than today, with an average MAT anomaly of −5.3 °C relative to present-day conditions and more pronounced changes in MTCO than in MTWA.
-
They suggest a gradual warming on average from the LGM to the Late Glacial. Nonetheless, the spatial pattern of temperature change between these two periods was highly heterogeneous across Europe, with a faster increase in temperature in the northwestern parts of the continent than in the southwestern and eastern regions.
-
They produce spatial temperature patterns that are more different than those derived from GCMs, although these divergences are generally included in the range of GCM uncertainties, which points to the relevance of using paleoclimatic approaches based on multiple proxies.
Future work in under-sampled regions, such as the Balkans, North Africa or the Middle East, is expected to enhance the robustness of our conclusions. This could allow for the integration of rodent-based inferences as input or comparison data for GCMs and for new comprehensive analyses including other additional proxies. A similar approach including other taxa such as Eulipotyphla or large mammals could also be developed to explore, in particular, other climatic variables such as precipitation or aridity. These initiatives should further improve our knowledge of past climate changes.
All the analyses of the paper were performed using the R software (R core team, 2023). The Bioclimatic Analysis is freely available through the PalBER R scripts at https://github.com/AurelienRoyer/PalBER (last access: 15 October 2025). All the R scripts generated for all calculations, as well as the modified functions, and data are available at https://github.com/AurelienRoyer/Climate-spatial-Interpolations-LGM-LG (last access: 15 October 2025) and https://doi.org/10.5281/zenodo.14905209 (Royer, 2025) in order to be fully reproducible.
The supplement related to this article is available online at https://doi.org/10.5194/cp-21-1821-2025-supplement.
AR designed the study. BL, IM and MHF provided the data on the modern rodent assemblages and Climatic Restriction Index of rodent species. JC and BP provided GCMs and ERA5 data. AR provided rodent fossil data. AR, RL and JC developed the methodology. AR wrote R scripts. AR, JC, RL, SB, BL, IM, BP, SM and MHF participated in analysing and writing and provided edits and comments on subsequent drafts.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
This research was partially financed by PID2022-138275NB-I00 projects (Ministry of Science and Innovation, Spain); Sara Gamboa was funded by the Ministry of Universities and the Next Generation European Union programme through a Margarita Salas Grant from Universidad Complutense de Madrid (CT31/21) and by a Fundación Ramón Areces Postdoctoral Fellowship (2025). Iris Menéndez was funded by the Alexander von Humboldt Foundation through a Humboldt Postdoctoral Fellowship.
This research was partially financed by PID2022-138275NB-I00 projects (Ministry of Science and Innovation, Spain); Sara Gamboa was funded by the Ministry of Universities and the Next Generation European Union programme through a Margarita Salas Grant from Universidad Complutense de Madrid (CT31/21). Iris Menéndez was funded by the Alexander von Humboldt Foundation through a Humboldt Postdoctoral Fellowship. Aurélien Royer was financed by Biogéosciences laboratory resources.
This paper was edited by Claudio Latorre and reviewed by Juan Manuel Lopez and one anonymous referee.
Aguilar, J. P., Pélissié, T., Sigé, B., and Michaux, J.: Occurrence of the stripe field mouse lineage (Apodemus agrarius Pallas 1771; Rodentia; Mammalia) in the Late Pleistocene of southwestern France, C. R.-Palevol., 7, 217–225, https://doi.org/10.1016/j.crpv.2008.02.004, 2008.
Albouy, B., Paquin, S., Riel-Salvatore, J., Kageyama, M., Vrac, M., and Burke, A.: Evaluating the impact of climate change and millennial variability on the last Neanderthal populations in Europe (Marine Isotope Stage 3), Quaternary Sci. Rev., 338, 108812, https://doi.org/10.1016/j.quascirev.2024.108812, 2024.
Allen, R., Siegert, M. J., and Payne, A. J.: Reconstructing glacier-based climates of LGM Europe and Russia – Part 3: Comparison with previous climate reconstructions, Clim. Past, 4, 265–280, https://doi.org/10.5194/cp-4-265-2008, 2008.
Allen, J. R., Hickler, T., Singarayer, J. S., Sykes, M. T., Valdes, P. J., and Huntley, B.: Last glacial vegetation of northern Eurasia, Quaternary Sci. Rev., 29, 2604–2618, https://doi.org/10.1016/j.quascirev.2010.05.031, 2010.
Álvarez-Vena, A., Álvarez-Lao, D. J., Laplana, C., Quesada, J. M., Rojo, J., Garcia-Sanchez, E., and Menendez, M.: Environmental context for the Late Pleistocene (MIS 3) transition from Neanderthals to early Modern Humans: analysis of small mammals from La Güelga Cave, Asturias, northern Spain, Palaeogeogr. Palaeoclimatol. Palaeoecol., 562, 110096, https://doi.org/10.1016/j.palaeo.2020.110096, 2021.
Arbez, L., Royer, A., Schreve, D., Laffont, R., David, S., and Montuire, S.: The missing Myopus: plugging the gaps in Late Pleistocene small mammal identification in western Europe with geometric morphometrics, J. Quaternary Sci., 36, 224–238, https://doi.org/10.1002/jqs.3269, 2021.
Arbez, L., Tereza, H., Royer, A., Sophie, M., and Ivan, H.: The wood lemming and the development of taiga in Late Pleistocene Central Europe, Quaternary Sci. Rev., 303, 107974, https://doi.org/10.1016/j.quascirev.2023.107974, 2023.
Argus, D. F., Peltier, W. R., Drummond, R., and Moore, A. W.: The Antarctica component of postglacial rebound model ICE-6G_C (VM5a) based upon GPS positioning, exposure age dating of ice thicknesses, and relative sea level histories, Geophys. J. Int., 198, 537–563, https://doi.org/10.1093/gji/ggu140, 2014.
Avery, D. M.: Late pleistocene coastal environment of the Southern Cape province of South Africa: Micromammals from Klasies river Mouth, J. Archaeol. Sci., 14, 405–421, https://doi.org/10.1016/0305-4403(87)90028-8, 1987.
Baca, M., Nadachowski, A., Lipecki, G., Mackiewicz, P., Marciszak, A., Popović, D., Socha, P. Stefaniak, K., and Wojtal, P.: Impact of climatic changes in the Late Pleistocene on migrations and extinctions of mammals in Europe: four case studies, Geol. Q., 61, 291–304, 2017.
Baca, M., Popović, D., Agadzhanyan, A. K., Baca, K., Conard, N. J., Fewlass, H., Filek, T., Golubiński, M., Horáček, I., Knul, M. V., Krajcarz, M., Krokhaleva, M., Lebreton, L., Lemanik, A., Maul, L. C., Nagel, D., Noiret, P., Primault, J., Rekovets, L., Rhodes, S., Royer, A., Serdyuk, N.V, Soressi, M., Stewart, J., Strukova, T., Talamo, S., Wilczyński, J., and Nadachowski, A.: Ancient DNA of narrow-headed vole reveal common features of the Late Pleistocene population dynamics in cold-adapted small mammals, Proc. R. Soc. B, 290, 20222238, https://doi.org/10.1098/rspb.2022.2238, 2023a.
Baca, M., Popović, D., Lemanik, A., Bañuls-Cardona, S., Conard, N. J., Cuenca-Bescós, G., Desclaux, E., Fewlass, H., Garcia, J. T., Hadravova, T., Heckel, G., Horáček, I., Knul, M. V., Lebreton, L., López-García, J. M., Luzi, E., Marković, Z., Mauch Lenardić, J., Murelaga, X., Petculescu, A., Popov, V., Putalova, T., Rhodes, S. E., Ridush, B., Royer, A., Stewart, J. R., Stojak, J., Talamo, S., Wang, X., Wójcik, J. M., and Nadachowski, A.: Ancient DNA reveals interstadials as a driver of common vole population dynamics during the last glacial period, J. Biogeogr., 50, 183–196, https://doi.org/10.1111/jbi.14521, 2023b.
Bartlein, P. J., Harrison, S. P., Brewer, S., Connor, S., Davis, B. A. S., Gajewski, K., Guiot, J., Harrison-Prentice, T. I., Henderson, A., Peyron, O., Scholze, M., Seppä, H., Shuman, B., Sugita, S., Thompson, R. S., Viau, A. E., Williams, J., and Wu, H.: Pollen-based continental climate reconstructions at 6 and 21 ka: a global synthesis, Clim. Dynam., 37, 775–802, https://doi.org/10.1007/s00382-010-0904-1, 2011.
Benjamin, J., Rovere, A., Fontana, A., Furlani, S., Vacchi, M., Inglis, R. H., Galili, E., Antonioli, F., Sivan, D., Miko, S., Mourtzas, N., Felja, I., Meredith-Williams, M., Goodman-Tchernov, B., Kolaiti, E., Anzidei, M., and Gehrels, R.: Late Quaternary sea-level changes and early human societies in the central and eastern Mediterranean Basin: An interdisciplinary review, Quatern. Int., 449, 29–57, https://doi.org/10.1016/j.quaint.2017.06.025, 2017.
Berto, C., Boscato, P., Boschin, F., Luzi, E., and Ronchitelli, A.: Paleoenvironmental and paleoclimatic context during the Upper Palaeolithic (late Upper Pleistocene) in the Italian Peninsula. The small mammal record from Grotta Paglicci (Rignano Garganico, Foggia, Southern Italy), Quaternary Sci. Rev., 168, 30–41, https://doi.org/10.1016/j.quascirev.2017.05.004, 2017.
Berto, C., López-García, J. M., and Luzi, E.: Changes in the Late Pleistocene small-mammal distribution in the Italian Peninsula, Quaternary Sci. Rev., 225, 106019, https://doi.org/10.1016/j.quascirev.2019.106019, 2019a.
Berto, C., Santaniello, F., and Grimaldi, S.: Palaeoenvironment and palaeoclimate in the western Liguria region (northwestern Italy) during the Last Glacial. The small mammal sequence of Riparo Mochi (Balzi Rossi, Ventimiglia), C. R.-Palevol., 18, 13–23, https://doi.org/10.1016/j.crpv.2018.04.007, 2019b.
Beyer, R. M., Krapp, M., and Manica, A.: High-resolution terrestrial climate, bioclimate and vegetation for the last 120,000 years, Sci. Data, 7, 236, https://doi.org/10.1038/s41597-020-0552-1, 2020.
Binney, H., Edwards, M., Macias-Fauria, M., Lozhkin, A., Anderson, P., Kaplan, J. O., Andreev, A., Bezrukova, E., Blyakharchuk, T., Jankovska, V., Khazina, I., Krivonogov, S., Kremenetski, K., Nield, K., Novenko, E., Ryabogina, N., Solovieva, N., Willis, K., and Zernitskaya, V.: Vegetation of Eurasia from the last glacial maximum to present: Key biogeographic patterns, Quaternary Sci. Rev., 157, 80–97, https://doi.org/10.1016/j.quascirev.2016.11.022, 2017.
Björck, S., Walker, M., Cwynar, L., Johnsen, S., Knudsen, K. L., Lowe, J., and Wohlfarth, B.: An event stratigraphy for the Last Termination in the North Atlantic region based on the Greenland ice-core record: a proposal by the INTIMATE group, J. Quaternary Sci., 13/4, 283–292, https://doi.org/10.1002/(SICI)1099-1417(199807/08)13:4<283::AID-JQS386>3.0.CO;2-A, 1998.
Brace, S., Ruddy, M., Miller, R., Schreve, D. C., Stewart, J. R., and Barnes, I.: The colonization history of British water vole (Arvicola amphibius (Linnaeus, 1758)): origins and development of the Celtic fringe, Proc. R. Soc. B: Biol. Sci., 283, 20160130, https://doi.org/10.1098/rspb.2016.0130, 2016.
Braconnot, P., Harrison, S. P., Kageyama, M., Bartlein, P. J., Masson-Delmotte, V., Abe-Ouchi, A., Otto-Bliesner, B., and Zhao, Y.: Evaluation of climate models using palaeoclimatic data, Nat. Clim. Change, 2, 417–424, https://doi.org/10.1038/nclimate1456, 2012.
Budich, R., Giorgetta, M., Jungclaus, J. H., and Reick, C. H.: The MPI-M Millennium Earth System Model: An Assembling Guide for the COSMOS Configuration, Tech. rep., 2010.
Burke, A., Peros, M. C., Wren, C. D., Pausata, F. S., Riel-Salvatore, J., Moine, O., de Vernal, A., Kageyama, M., and Boisard, S.: The archaeology of climate change: The case for cultural diversity, P. Natl. Acad. Sci. USA, 118, e2108537118, https://doi.org/10.1073/pnas.2108537118, 2021.
Calandra, I, Labonne, G, Mathieu, O, Henttonen, H, Lévêque, J, Milloux, M-J, Renvoisé, É, Montuire, S, and Navarro, N.: Isotopic partitioning by small mammals in the subnivium, Ecol. Evol., 5, 4132–4140, https://doi.org/10.1002/ece3.1653, 2015.
Carré, M., Braconnot, P., Elliot, M., D'agostino, R., Schurer, A., Shi, X., Marti, O., Lohmann, G., Jungclauss, J., Cheddadi, R., Abdelkader di Carlo, I., Cardich, J., Ochoa, D., Salas Gismondi, R., Pérez, A., Romero, P. E., Turcq, B., Corrège, T., and Harrison, S. P.: High-resolution marine data and transient simulations support orbital forcing of ENSO amplitude since the mid-Holocene, Quaternary Sci. Rev., 268, 107125, https://doi.org/10.1016/j.quascirev.2021.107125, 2021.
Ceregatti, L., Berto, C., Fewlass, H., Baca, M., Luzi, E., Brancaleoni, G., Pereswiet-Soltan, A., and Peresani, M.: Integration of direct radiocarbon dating, genetic studies and taxonomy of small mammals to investigate the chronology of past climatic oscillations: The Last Glacial Maximum sequence of Grotta della Ferrovia (Fabriano, Italy), Quaternary Sci. Rev., 309, 108095, https://doi.org/10.1016/j.quascirev.2023.108095, 2023.
Chaline, J.: Les rongeurs du pléistocène moyen et supérieur de France: (systématique, biostratigraphie, paléoclimatologie), Éditions du Centre national de la recherche scientifique, 403 p., ISBN: 2222014883, 1972.
Chaline, J. and Brochet, G.: Les Rongeurs: leurs significations paléoécologiques et paléoclimatiques in La Baume de Gigny (Jura), Gallia Préhistoire, Suppl. 27, 97–109, ISBN 2-222-04345-X, http://www.persee.fr/doc/galip_0072-0100_1989_sup_27_1_2563 (last access: 15 October 205), 1989.
Comas-Bru, L., Harrison, S. P., Werner, M., Rehfeld, K., Scroxton, N., Veiga-Pires, C., and SISAL working group members: Evaluating model outputs using integrated global speleothem records of climate change since the last glacial, Clim. Past, 15, 1557–1579, https://doi.org/10.5194/cp-15-1557-2019, 2019.
Cordy, J. M.: Palaeoecology of the Late Glacial and early Postglacial of Belgium and neighbouring areas, in: The Late Glacial in Northwest Europe: Human adaptation and environmental change at the end of the Pleistocene, edited by: Barton, N., Roberts, A. J., and Roe, D. A., Council for British Archaeology, 40–47, ISBN 9781872414157, ISBN 187241415X, 1991.
Courtiol, A. and Rousset, F.: Modelling isoscapes using mixed models, bioRxiv [preprint], 207662, https://doi.org/10.1101/207662, 23 October 2017.
Courtiol, A., Rousset, F., Rohwäder, M. S., Soto, D. X., Lehnert, L. S., Voigt, C. C., Hobson, K. A., Wassenaar, L. I., and Kramer-Schadt, S.: Isoscape computation and inference of spatial origins with mixed models using the R package IsoriX, in: Tracking animal migration with stable isotopes, Academic Press, 207–236, https://doi.org/10.1016/B978-0-12-814723-8.00009-X, 2019.
Crégut-Bonnoure, E., Boulbes, N., Desclaux, E., and Marciszak, A.: New insights into the LGM and LG in Southern France (Vaucluse): the mustelids, micromammals and horses from Coulet des Roches, Quaternary, 1, 19, https://doi.org/10.3390/quat1030019, 2018.
Cuenca-Bescós, G., Straus, L. G., Morales, M. R. G., and Pimienta, J. C. G.: The reconstruction of past environments through small mammals: from the Mousterian to the Bronze Age in El Mirón Cave (Cantabria, Spain), J. Archaeol. Sci., 36, 947–955, https://doi.org/10.1016/j.jas.2008.09.025., 2009.
Cuenca-Bescós, G., Rofes, J., López-García, J. M., Blain, H. A., Roger, J., Galindo-Pellicena, M. A., Bennásar-Serra, M. L., Melero-Rubio, M., Arsuaga, J. L., Bermúdez de Castro, J. M., and Carbonell, E.: Biochronology of Spanish Quaternary small vertebrate faunas, Quatern. Int., 212, 109–119, https://doi.org/10.1016/j.quaint.2009.06.007, 2010.
Cyrek, K., Nadachowski, A., Madeyska, T., Bochenski, Z., Tomek, T., Wojtal, P., Miekina, B., Lipecki, G., Garapich, A., Rzebik-Kowalska, B., Stworzewicz, E.,Wolsan, M., Godawa, J., Kosciów, R., Fostowicz-Frelik, L., and Szyndlar, Z.: Excavation in the Deszczowa Cave (Kroczyckie Rocks, Czestochowa Upland, Central Poland), Folia Quaternaria, 71, 5–84, 2000.
Daams, R. and van der Meulen, A. J.: Paleoenvironmental and paleoclimatic interpretation of micromammal faunal successions in the Upper Oligocene and Miocene of North Central Spain, Paleobiol. Cont., 14, 241–257, 1984.
Daams, R., Freudenthal, M., and van der Meulen, A. J.: Ecostratigraphy of micromammal faunas from the Neogene of Spain, Scr. Geol., Special Issue, 1, 287–302, 1988.
Danukalova, G. and Yakovlev, A.: A review of biostratigraphical investigations of palaeolithic localities in the Southern Urals region, Quatern. Int., 149, 37–43, https://doi.org/10.1016/j.quaint.2005.11.016, 2006.
Davis, B. A. S., Fasel, M., Kaplan, J. O., Russo, E., and Burke, A.: The climate and vegetation of Europe, northern Africa, and the Middle East during the Last Glacial Maximum (21 000 yr BP) based on pollen data, Clim. Past, 20, 1939–1988, https://doi.org/10.5194/cp-20-1939-2024, 2024.
Discamps, E., Thomas, M., Dancette, C., Gravina, B., Plutniak, S., Royer, A., Angelin, A., Bachellerie, F., Beauval, C., Bordes, J.-G., Deschamps, M., Ducasse, S., Langlais, M., Laroulandie, V., Mallye, J.-B., Michel, A., Perrin, T., and Rendu, W.: Breaking free from field layers: the interest of Post-Excavation Stratigraphies (PES) for producing reliable archaeological interpretations and increasing chronological resolution, J. Paleolit. Archaeol., 6, 29, https://doi.org/10.1007/s41982-023-00155-x, 2023.
Domínguez-García, Á. C., Laplana, C., Sevilla, P., Álvarez-Vena, A., and Collado Giraldo, H.: Small mammals of the Holocene sequence of Postes Cave (SW Spain): biogeographic and palaeoenvironmental implications for southwestern Iberia, Hist. Biol., 35, 483–497, https://doi.org/10.1080/08912963.2022.2045981, 2023.
Domínguez-García, Á.C., López-García, J. M., Núñez-Lahuerta, C., Galán, J., and Cuenca-Bescós, G.: Palaeoclimatic analysis of Quaternary terrestrial small mammal assemblages from the Sierra de Atapuerca (Burgos, Spain), Palaeogeogr. Palaeoclimatol. Palaeoecol., 112532, https://doi.org/10.1016/j.palaeo.2024.112532, 2024.
Dufresne, J. L., Foujols, M. A., Denvil, S., Caubel, A., Marti, O., Aumont, O., Balkanski, Y., Bekki, S., Bellenger, H., Benshila, R., Bony, S., Braconnot, P., Brockmann, P., Cadule, P., Cheruy, F., Codron, F., Cozic, A., Cugnet, D., de Noblet, N., Duvel, J.-P., Ethé, C., Fairhead, L., Fichefet, T., Flavoni, S., Friedlingstein, P., Grandpeux, J.-Y., Guez, L., Guilyardi, E., Hauglustaine, D., Hourdin, F., Idelkadi, A., Ghattas, J., Joussaume, S., Kageyama, M., Krinner, G., Labetoulle, S., Lahellec, A., Lefebvre, M.-P., Lefevre, F., Levy, C., Li, Z. X., Lloyd, J., Lott, F., Madec, G., Mancip, M., Marchand, M., Masson, S., Meurdesoif, Y., Mignot, J., Musat, I., Parouty, S., Polcher, J., Rio, C., Schulz, M., Swingedouw, D., Szopa, S., Talandier,C., Terray, P., Viovy, N., and Vuichard, N.: Climate change projections using the IPSL-CM5 Earth System Model: from CMIP3 to CMIP5, Clim. Dynam., 40, 2123–2165, https://doi.org/10.1007/s00382-012-1636-1, 2013.
Emile-Geay, J., Cobb, K. M., Carré, M., Braconnot, P., Leloup, J., Zhou, Y., Harrison, S. P., Corrège, T., McGregor, H. V., Collins, M., Discoll, R., Elliot, M., Schneifer, B., and Tudhope, A.: Links between tropical Pacific seasonal, interannual and orbital variability during the Holocene, Nat. Geosci., 9, 168–173, https://doi.org/10.1038/ngeo2608, 2016.
Erbajeva, M. and Alexeeva, N.: Chapter 21. Late Cenozoic Mammal Faunas of the Baikalian Region: Composition, Biochronology, Dispersal, and Correlation with Central Asia, in: Fossil mammals of Asia: Neogene biostratigraphy and chronology, Columbia University Press, https://doi.org/10.7312/wang15012-021, 495–507, 2013.
Escudé, É., Renvoisé, É., Lhomme, V., and Montuire, S.: Why all vole molars (Arvicolinae, Rodentia) are informative to be considered as proxy for Quaternary paleoenvironmental reconstructions, J. Archaeol. Sci., 40, 11–23, https://doi.org/10.1016/j.jas.2012.03.003, 2013.
Eskelinen, O.: Diet of the wood lemming Myopus schisticolor, Ann. Zool. Fenn., 39, 49–57, http://www.jstor.org/stable/23735802, 2002.
Fagoaga, A., Blain, H. A., Marquina-Blasco, R., Laplana, C., Sillero, N., Hernández, C. M., Mallol, C., Galván, B., and Ruiz-Sánchez, F. J.: Improving the accuracy of small vertebrate-based palaeoclimatic reconstructions derived from the Mutual Ecogeographic Range. A case study using geographic information systems and UDA-ODA discrimination methodology, Quaternary Sci. Rev., 223, 105969, https://doi.org/10.1016/j.quascirev.2019.105969, 2019.
FAUNMAP Working Group: Spatial response of mammals to Late Quaternary environmental fluctuations, Science, 272, 1601–1606, https://doi.org/10.1126/science.272.5268.1601, 1996.
Fernández-García, M., López-García, J. M., and Lorenzo, C.: Palaeoecological implications of rodents as proxies for the Late Pleistocene–Holocene environmental and climatic changes in northeastern Iberia, C. R.-Palevol, 15, 707–719, https://doi.org/10.1016/j.crpv.2015.08.005, 2016.
Fernández-García, M., López-García, J. M., Royer, A., Lécuyer, C., Allué, E., Burjachs, F., Chacón, G. M., Saladié, P., Vallverdú, J., and Carbonell, E.: Combined palaeoecological methods using small-mammal assemblages to decipher environmental context of a long-term Neanderthal settlement in northeastern Iberia, Quaternary Sci. Rev., 228, 106072, https://doi.org/10.1016/j.quascirev.2019.106072, 2020.
Flynn, L. J., Morgan, M. E., Barry, J. C., Raza, S. M., Cheema, I. U., and Pilbeam, D.: Siwalik rodent assemblages for NOW: Biostratigraphic resolution in the Neogene of South Asia, in: Evolution of Cenozoic Land Mammal Faunas and Ecosystems: 25 Years of the NOW Database of Fossil Mammals, Springer International Publishing, Cham, 43–58, https://doi.org/10.1007/978-3-031-17491-9_4, 2023.
Furió, M., Casanovas-Vilar, I., and Van Den Hoek Ostende, L. W.: Predictable structure of Miocene insectivore (Lipotyphla) faunas in Western Europe along a latitudinal gradient, Palaeogeogr. Palaeoclimatol. Palaeoecol., 304, 219–229, https://doi.org/10.1016/j.palaeo.2010.01.039, 2011.
García-Alix, A., Minwer-Barakat, R., Martín Suárez, E., Freudenthal, M., and Martín, J. M.: Late Miocene–Early Pliocene climatic evolution of the Granada Basin (southern Spain) deduced from the paleoecology of the micromammal associations, Palaeogeogr. Palaeoclimatol. Palaeoecol., 265, 214–225, https://doi.org/10.1016/j.palaeo.2008.04.005, 2008.
Garcia-Ibaibarriaga, N., Suárez-Bilbao, A., Iriarte-Chiapusso, M. J., Arrizabalaga, A., and Murelaga, X.: Palaeoenvironmental dynamics in the Cantabrian Region during Greenland stadial 2 approached through pollen and micromammal records: State of the art, Quatern. Int., 506, 14–24, https://doi.org/10.1016/j.quaint.2018.12.004, 2019.
García-Morato, S., Domínguez-García, Á. C., Sevilla, P., Laplana, C., and Fernández-Jalvo, Y.: The last 20,000 years of climate change in the Iberian Peninsula characterized by the small-mammal assemblages, Palaeogeogr. Palaeoclimatol. Palaeoecol., 655, 112545, https://doi.org/10.1016/j.palaeo.2024.112545, 2024.
Gent, P. R., Danabasoglu, G., Donner, L. J., Holland, M. M., Hunke, E. C., Jayne, S.R, Lawrence, D. M., Neale, R. B., Rasch, P. J., Vertenstein, M., Worley, P. H., Yang, Z.-L., and Zhang, M.: The community climate system model version 4, J. Climate, 24, 4973–4991, https://doi.org/10.1175/2011JCLI4083.1, 2011.
Giorgetta, M. A., Jungclaus, J., Reick, C. H., Legutke, S., Bader, J., Böttinger, M., Brovkin, V., Crueger, T., Esch, M., Fieg, K., Glushak, K., Gayler, V., Haak, H., Hollweg, H.-D., Ilyina, T.,Kinne, S., Kornblueh, L., Matei, D., Mauritsen, T., Mikolajewicz, U., Mueller, W., Notz, D., Pithan, F., Raddatz, T., Rast, S., Redler, R., Roeckner, E, Schmidt, H., Schnur, R., Segschneider, J., Six, K. D., Stockhause, M., Timmreck, C., Wegner, J., Widmann, H., Wieners, K.-H., Claussen, M., Marotzke, J., and Stevens, B.: Climate and carbon cycle changes from 1850 to 2100 in MPI-ESM simulations for the Coupled Model Intercomparison Project phase 5, JAMES, 5, 572–597, https://doi.org/10.1002/jame.20038, 2013.
González-Varo, J. P.: Avian seed dispersal out of the forests: A view through the lens of Pleistocene landscapes, J. Ecol., 113, 510–517, https://doi.org/10.1111/1365-2745.14457, 2025.
Graham, R. W.: Quaternary Mammal Communities: Relevance of the Individualistic Response and Non-Analogue Faunas, Pal. Soc. Papers, 11, 141–158, https://doi.org/10.1017/S1089332600001297, 2005.
Hanquet, C., Desclaux, E., and Hinguant, S.: Les rongeurs des niveaux solutréens de la grotte Rochefort (Saint-Pierre-sur-Erve, Mayenne, France): un référentiel inédit pour le dernier maximum glaciaire du nord-ouest de la France, Quaternaire, 27, 341–352, https://doi.org/10.4000/quaternaire.7735, 2016.
Harzhauser, M., Daxner-Höck, G., Erbajeva, M. A., López-Guerrero, P., Maridet, O., Oliver, A., Piller, W. E., Göhlich, U., and Ziegler, R.: Oligocene and early Miocene mammal biostratigraphy of the Valley of Lakes in Mongolia, Paleobiodivers. Paleoenviron., 97, 219–231, https://doi.org/10.1007/s12549-016-0264-x, 2017.
Haywood, A. M., Chandler, M. A., Valdes, P. J., Salzmann, U., Lunt, D. J., and Dowsett, H. J.: Comparison of mid-Pliocene climate predictions produced by the HadAM3 and GCMAM3 General Circulation Models, Glob. Planet. Change, 66, 208–224, https://doi.org/10.1016/j.gloplacha.2008.12.014, 2009.
Henrot, A. J., Utescher, T., Erdei, B., Dury, M., Hamon, N., Ramstein, G., Krapp, M., Herold, N., Goldner, A., E. Favre, E., Munhoven, G., and François, L.: Middle Miocene climate and vegetation models and their validation with proxy data, Palaeogeogr. Palaeoclimatol. Palaeoecol., 467, 95–119, https://doi.org/10.1016/j.palaeo.2016.05.026, 2017.
Hersbach, H., Bell, B., Berrisford, P., Hirahara, S., Horányi, A., Muñoz-Sabater, J., Nicolas, J., Peubey, C., Radu, R., Schepers, D., Simmons, A., Soci, C., Abdalla, S., Abellan, X., Balsamo, G., Bechtold, P., Biavati, G., Bidlot, J., Bonavita, M., De Chiara, G., Dahlgren, P., Dee, D., Diamantakis, M., Dragani, R., Flemming, J., Forbes, R., Fuentes M., Geer, A., Haimberger, L., Healy, S., Hogan, R. J., Hólm, E., Janisková, M., Keely, S., Laloyaux, P., Lopez, P., Lupu, C., Radnoti, G., de Rosnay, P., Rozum, I., Vamborg, F., Villaume, S., and Thépaut, J. N.: The ERA5 global reanalysis, Q. J. Roy. Meteorol. Soc., 146, 1999–2049, https://doi.org/10.1002/qj.3803, 2020.
Hernández Fernández, M.: Bioclimatic discriminant capacity of terrestrial mammal faunas, Glob. Ecol. Biogeogr., 10, 189–204, https://doi.org/10.1046/j.1466-822x.2001.00218.x, 2001.
Hernández Fernández, M.: Rodent paleofaunas as indicators of climatic change in Europe during the last 125,000 years, Quaternary Res., 65, 308–323, https://doi.org/10.1016/j.yqres.2005.08.022, 2006.
Hernández Fernández, M. and Peláez-Campomanes, P.: The bioclimatic model: a method of palaeoclimatic qualitative inference based on mammal associations, Glob. Ecol. Biogeogr., 12, 507–517, https://doi.org/10.1046/j.1466-822X.2003.00057.x, 2003.
Hernández Fernández, M. and Peláez-Campomanes, P.: Quantitative palaeoclimatic inference based on terrestrial mammal faunas, Glob. Ecol. Biogeogr., 39–56, https://doi.org/10.1111/j.1466-822X.2004.00125.x, 2005.
Hernández Fernández, M., Álvarez Sierra, M.Á., and Peláez-Campomanes, P.: Bioclimatic analysis of rodent palaeofaunas reveals severe climatic changes in Southwestern Europe during the Plio-Pleistocene, Palaeogeogr. Palaeoclimatol. Palaeoecol., 251, 500–526, https://doi.org/10.1016/j.palaeo.2007.04.015, 2007.
Hijmans, R. J., Bivand, R., Forner, K., Ooms, J., Pebesma, E., and Sumner, M. D.: Package “terra”, Maintainer: Vienna, Austria [code], https://rspatial.github.io/terra/ (last access: 15 October 2025), 2022.
Hinton, M. A.: Monograph of the voles and lemmings, British Museum (Natural History), London, https://doi.org/10.5962/bhl.title.8319, 1926.
Hinz, M., Schmid, C., Knitter, D., and Tietze, C.: oxcAAR: Interface to 'OxCal' Radiocarbon Calibration [code], https://doi.org/10.7892/boris.127058, 2018.
Hokr, Z.: A method of the quantitative determination of the climate in the Quaternary period by means of mammal associations, Sbornik of the geological Survey of Czechslovakia, 18, 209–219, 1951.
IUCN: The IUCN Red List of Threatened Species, Version 3, May 2017, https://www.iucnredlist.org (last access: 23 June 2021), 2021.
Izvarin, E., Ulitko, A., Panina, S., Zazovskaya, E., and Nekrasov, A.: Voronin Grotto (Middle Urals, Russia): Analysis of vertebrate assemblage with taphonomic remarks and reconstruction of the Late Bronze Age and Early Iron Age human environment in the east end of Europe based on small mammals, Quatern. Int., 632, 178–191, https://doi.org/10.1016/j.quaint.2022.02.031, 2022.
Jackson, S. T. and Overpeck, J. T.: Responses of plant populations and communities to environmental changes of the Late Quaternary, Paleobiology, 26, 194–220, https://doi.org/10.1017/S0094837300026932, 2000.
Jeannet, M.: L'environnement tardiglaciaire préalpin: essai de restitution basée sur le potentiel climatique et écologique des microvertébrés, Revue archéologique de l'Est, 58, 5–56, 2009.
Jeannet, M.: L'écologie quantifiée. Essai de description de l'environnement continental à l'aide des microvertébrés, Préhistoires méditerranéennes, 1, https://doi.org/10.4000/pm.492, 2010.
Jeannet, M.: Vergisson II (Saône-et-Loire, France): microfaune et environnement de l'Homme de Néandertal, Quaternaire, 29, 233–242, https://doi.org/10.4000/quaternaire.10270, 2018.
Johannsen, F., Ermida, S., Martins, J. P., Trigo, I. F., Nogueira, M., and Dutra, E.: Cold bias of ERA5 summertime daily maximum land surface temperature over Iberian Peninsula, Remote Sens., 11, 2570, https://doi.org/10.3390/rs11212570, 2019.
Jost, A., Lunt, D., Kageyama, M., Abe-Ouchi, A., Peyron, O., Valdez, P. J., and Ramstein, G.: High-resolution simulations of the last glacial maximum climate over Europe: a solution to discrepancies with continental palaeoclimatic reconstructions?, Clim. Dynam., 24, 577–590, https://doi.org/10.1007/s00382-005-0009-4, 2005.
Jovanović, M., Bogićević, K., Nenadić, D., Agustí, J., Sánchez-Bandera, C., López-García, J. M., and Blain, H. A.: New paleoecological perspectives on Late Pleistocene Neanderthals in northern Balkans: the rodent assemblages from Smolućka cave (Serbia), Archaeol. Anthropol. Sci., 14, 169, https://doi.org/10.1007/s12520-022-01624-0, 2022.
Kageyama, M. and Paillard, D.: Modeling and Paleoclimatology, Paleoclimatology, 319–342, https://doi.org/10.1007/978-3-030-24982-3_25, 2021.
Kageyama, M., Peyron, O., Pinot, S., Tarasov, P., Guiot, J., Joussaume, S., and Ramstein, G.: The Last Glacial Maximum climate over Europe and western Siberia: a PMIP comparison between models and data, Clim. Dynam., 17, 23–43, https://doi.org/10.1007/s003820000095, 2001.
Kageyama, M., Sime, L. C., Sicard, M., Guarino, M.-V., de Vernal, A., Stein, R., Schroeder, D., Malmierca-Vallet, I., Abe-Ouchi, A., Bitz, C., Braconnot, P., Brady, E. C., Cao, J., Chamberlain, M. A., Feltham, D., Guo, C., LeGrande, A. N., Lohmann, G., Meissner, K. J., Menviel, L., Morozova, P., Nisancioglu, K. H., Otto-Bliesner, B. L., O'ishi, R., Ramos Buarque, S., Salas y Melia, D., Sherriff-Tadano, S., Stroeve, J., Shi, X., Sun, B., Tomas, R. A., Volodin, E., Yeung, N. K. H., Zhang, Q., Zhang, Z., Zheng, W., and Ziehn, T.: A multi-model CMIP6-PMIP4 study of Arctic sea ice at 127 ka: sea ice data compilation and model differences, Clim. Past, 17, 37–62, https://doi.org/10.5194/cp-17-37-2021, 2021a.
Kageyama, M., Harrison, S. P., Kapsch, M.-L., Lofverstrom, M., Lora, J. M., Mikolajewicz, U., Sherriff-Tadano, S., Vadsaria, T., Abe-Ouchi, A., Bouttes, N., Chandan, D., Gregoire, L. J., Ivanovic, R. F., Izumi, K., LeGrande, A. N., Lhardy, F., Lohmann, G., Morozova, P. A., Ohgaito, R., Paul, A., Peltier, W. R., Poulsen, C. J., Quiquet, A., Roche, D. M., Shi, X., Tierney, J. E., Valdes, P. J., Volodin, E., and Zhu, J.: The PMIP4 Last Glacial Maximum experiments: preliminary results and comparison with the PMIP3 simulations, Clim. Past, 17, 1065–1089, https://doi.org/10.5194/cp-17-1065-2021, 2021b.
Kalthoff, D. C.: The small mammals from the Kettig site (Rhineland-Palatinate, Germany) within the framework of the Alleröd-age mammalian fauna of central and southern Germany, Palaontol. Z., 72, 407–424, https://doi.org/10.1007/BF02988369, 1998.
Kosintsev, P. A. and Bachura, O. P.: Late Pleistocene and Holocene mammal fauna of the Southern Urals, Quatern. Int., 284, 161–170, https://doi.org/10.1016/j.quaint.2012.06.022, 2013.
Lambeck, K., Rouby, H., Purcell, A., Sun, Y., and Sambridge, M.: Sea level and global ice volumes from the Last Glacial Maximum to the Holocene, P. Natl. Acad. Sci. USA, 111, 15296–15303, https://doi.org/10.1073/pnas.1411762111, 2014.
Laplana, C., Muñoz Jiménez, A., Pueyo Morer, E. L., and Sevilla, P.: Los micromamíferos y la interpretación ambiental de los yacimientos de la Cuenca de Villarroya, in: Villarroya: yacimiento clave de la paleontología riojana, edited by: Alberdi, M. T., Azanza, B., and Cervantes, E., Instituto de Estudios Riojanos, Logroño, 281–296, ISBN 978-84-9960-093-2, 2016.
Latombe, G., Burke, A., Vrac, M., Levavasseur, G., Dumas, C., Kageyama, M., and Ramstein, G.: Comparison of spatial downscaling methods of general circulation model results to study climate variability during the Last Glacial Maximum, Geosci. Model Dev., 11, 2563–2579, https://doi.org/10.5194/gmd-11-2563-2018, 2018.
Lebreton, L. and López-García, J. M.: Stage or Sub-Stage: The Contribution of Small Mammals to the Characterization of Middle Pleistocene Local Climate Variation, Quaternary, 6, 54, https://doi.org/10.3390/quat6040054, 2023.
Legendre, S., Montuire, S., Maridet, O., and Escarguel, G.: Rodents and climate: a new model for estimating past temperatures, Earth Planet. Sci. Lett., 235, 408–420, https://doi.org/10.1016/j.epsl.2005.04.018, 2005.
Lemanik, A., Baca, M., Wertz, K., Socha, P., Popović, D., Tomek, T., Lipecki, G., Kraszewska, A., Miękina, B., Żeromska, A., Pereswiet-Soltan, A., Szyndlar, Z., Cieśla, M., Valde-Nowak, P., Mackiewicz, P., and Nadachowski, A.: The impact of major warming at 14.7 ka on environmental changes and activity of Final Palaeolithic hunters at a local scale (Orawa-Nowy Targ Basin, Western Carpathians, Poland, Archaeol. Anthropol. Sci., 12, 66, https://doi.org/10.1007/s12520-020-01020-6, 2020.
Lenardić, J. M.: First record of Dicrostonyx (Rodentia, Mammalia) in the late Pleistocene? Holocene sediments of Croatia, Geol. Croat., 66, 183–189, https://doi.org/10.4154/GC.2013.15, 2013.
Lenardić, J. M., Sršen, A. O., and Radović, S.: Quaternary fauna of the Eastern Adriatic (Croatia) with the special review on the Late Pleistocene sites, Quatern. Int., 494, 130–151, https://doi.org/10.1016/j.quaint.2017.11.028, 2018.
Leonardi, M., Hallett, E. Y., Beyer, R., Krapp, M., and Manica, A.: pastclim 1.2: an R package to easily access and use paleoclimatic reconstructions, Ecography, 2023, e06481, https://doi.org/10.1111/ecog.06481, 2023.
Linchamps, P., Stoetzel, E., Robinet, F., Hanon, R., Latouche, P., and Cornette, R.: Bioclimatic inference based on mammal community using machine learning regression models: perspectives for paleoecological studies, Front. Ecol. Evol., 11, 1178379, https://doi.org/10.3389/fevo.2023.1178379, 2023.
López-García, J. M., Blain, H. A., Cuenca-Bescós, G., Ruiz-Zapata, M. B., Dorado-Valiño, M., Gil-García, M. J., Valdeolmillos, A., Ortega, A. I., Carretero, J. M., Arsuaga, J. L., Bermúdez de Castro, J. M., and Carbonell, E.: Palaeoenvironmental and palaeoclimatic reconstruction of the Latest Pleistocene of El Portalón Site, Sierra de Atapuerca, northwestern Spain, Palaeogeogr. Palaeoclimatol. Palaeoecol., 292, 453–464, https://doi.org/10.1016/j.palaeo.2010.04.006, 2010.
López-García, J. M., Luzi, E., and Peresani, M.: Middle to Late Pleistocene environmental and climatic reconstruction of the human occurrence at Grotta Maggiore di San Bernardino (Vicenza, Italy) through the small-mammal assemblage, Quaternary Sci. Rev., 168, 42–54, https://doi.org/10.1016/j.quascirev.2017.05.005, 2017.
López-García, J. M., Cuenca-Bescós, G., Galindo-Pellicena, M.Á., Luzi, E., Berto, C., Lebreton, L., and Desclaux, E.: Rodents as indicators of the climatic conditions during the Middle Pleistocene in the southwestern Mediterranean region: implications for the environment in which hominins lived, J. Hum. Evol., 150, 102911, https://doi.org/10.1016/j.jhevol.2020.102911, 2021.
López-García, J. M., Blain, H. A., Goffette, Q., Cousin, C., and Folie, A.: Environmental and climatic inferences for Marine Isotope Stage 2 of southern Belgium (Meuse valley, Namur Province) based on rodent assemblages, Palaeogeogr. Palaeoclimatol. Palaeoecol., 636, 111976, https://doi.org/10.1016/j.palaeo.2023.111976, 2024.
Lundelius, E. L.: The implications of disharmonious assemblages for Pleistocene extinctions, J. Archaeol. Sci., 16, 407–417, https://doi.org/10.1016/0305-4403(89)90015-0, 1989.
Luzi, E., Berto, C., Calattini, M., Tessaro, C., and Galiberti, A.: Non-analogue communities in the Italian Peninsula during Late Pleistocene: The case of Grotta del Sambuco, Quatern. Int., 632, 132–138, https://doi.org/10.1016/j.quaint.2022.02.007., 2022a.
Luzi, E., Blanco-Lapaz, À., Rhodes, S. E., and Conard, N. J.: Paleoclimatic and paleoenvironmental reconstructions based on the small vertebrates from the Middle Paleolithic of Hohle Fels Cave, SW Germany, Archaeol. Anthropol. Sci., 14, 107, https://doi.org/10.1007/s12520-022-01568-5, 2022b.
Lyman, R. L.: Quantitative paleozoology, Cambridge University Press, ISBN: 9780511813863, 2008.
Lyman, R. L.: Paleoenvironmental reconstruction from faunal remains: ecological basics and analytical assumptions, J. Archaeol. Res., 25, 315–371, https://doi.org/10.1007/s10814-017-9102-6, 2017.
Lyman, R. L.: Assumptions and protocol of the taxonomic identification of faunal remains in zooarchaeology: A North American perspective, J. Archaeol. Method Theory, 26, 1376–1438, https://doi.org/10.1007/s10816-019-09414-0., 2019.
Magyari, E. K., Gasparik, M., Major, I., Lengyel, G., Pál, I., Virág, A., Korponai, J., Haliuc, A., Szabó, Z., and Pazonyi, P.: Mammal extinction facilitated biome shift and human population change during the last glacial termination in East-Central Europe, Sci. Rep.-UK, 12, 6796, https://doi.org/10.1038/s41598-022-10714-x, 2022.
Malhi, Y., Doughty, C. E., Galetti, M., Smith, F. A., Svenning, J. C., and Terborgh, J. W.: Megafauna and ecosystem function from the Pleistocene to the Anthropocene, P. Natl. Acad. Sci. USA, 113, 838–846, https://doi.org/10.1073/pnas.1502540113, 2016.
Mansino, S., Ruiz-Sánchez, F. J., Fierro, I., and Montoya, P.: Mio-Pliocene rodent assemblages from Alcoi Forn (Alcoy Basin, Eastern Spain). Biostratigraphical and palaeoclimatical inferences, Hist. Biol., 28, 1050–1065, https://doi.org/10.1080/08912963.2015.1102238, 2016.
Markova, E., Malygin, V., Montuire, S., Nadachowski, A., Quéré, J. P., and Ochman, K.: Dental variation in sibling species Microtus arvalis and M. rossiaemeridionalis (Arvicolinae, Rodentia): between-species comparisons and geography of morphotype dental patterns, J. Mamm. Evol., 17, 121–139, https://doi.org/10.1007/s10914-009-9128-8, 2010.
Markova, A. K., van Kolfschoten, T., Bohncke, S. J. P., Kosintsev, P. A., Mol, J., Puzachenko, A. Y., Simakova, A. N., Smirnov, N. G., Verpoorte, A., and Golovachev, I. B.: Evolution of European ecosystems during Pleistocene–Holocene transition (24–8 kyr BP), Institute of Geography of Russian Academy of Sciences, 279, ISBN: 9785891187955, 2019.
Markwick, P. J.: The palaeogeographic and palaeoclimatic significance of climate proxies for data-model comparisons, in: Deep-Time Perspectives on Climate Change: Marrying the Signal from Computer Models and Biological Proxies, edited by: Williams, M., Haywood, A. M., Gregory, F. J., and Schimdt, D. N., The Micropalaeontological Society, Special Publications, The Geological Society, London, 251–312, https://doi.org/10.1144/TMS002.13, ISBN: 9781862392403, 2007.
Marquet, J.-C.: Paléoenvironnement et chronologie des sites du domaine Atlantique français d'âge Pléistocène moyen et supérieur d'après l'étude des rongeurs, Les Cahiers de la Claise, Supplément 2, Tours, French, ISSN 0766-4702, 1993.
Martin, R. A., Honey, J. G., and Peláez-Camponanes, P.: The Meade Basin rodent project: a progress report, Paludicola, 3, 1–32, 2000.
Minwer-Barakat, R., Garcia-Alix, A., Suárez, E. M., Freudenthal, M., and Viseras, C.: Micromammal biostratigraphy of the Upper Miocene to lowest Pleistocene continental deposits of the Guadix basin, southern Spain, Lethaia, 45, 594–614, https://doi.org/10.1111/j.1502-3931.2012.00324.x, 2012.
Monaghan, N. T.: Irish Quaternary vertebrates, Adv. Irish Quat. Stud., 255–291, https://doi.org/10.2991/978-94-6239-219-9_9, 2017.
Montuire, S. and Desclaux, E.: Palaeoecological analysis of mammalian faunas and environmental evolution in the South of France during the Pleistocene, Boreas, 26, 355–365, https://doi.org/10.1111/j.1502-3885.1997.tb00861.x, 1997.
Montuire, S. and Marcolini, F.: Palaeoenvironmental significance of the mammalian faunas of Italy since the Pliocene, J. Quaternary Sci., 17, 87–96, https://doi.org/10.1002/jqs.625, 2002.
Montuire, S., Michaux, J., Legendre, S., and Aguilar, J. P.: Rodents and climate. 1. A model for estimating past temperatures using arvicolids (Mammalia: Rodentia), Palaeogeogr. Palaeoclimatol. Palaeoecol., 128, 187–206, https://doi.org/10.1016/S0031-0182(96)00038-7, 1997.
Nadachowski, A.: Late Quaternary rodents of Poland with special reference to morphotype dentition analysis of voles, Panstwowe Wydawnictwo Naukowe, Warszawa – Krakow, 109 pp., ISBN 8301040033, ISBN 9788301040031, 1982.
Nadachowski, A., Valde-Nowak, P.: New Late Pleistocene faunal assemblages from Podhale Basin, Western Carpathians, Poland: preliminary results, Acta Zool. Crac., 58, 181–194, https://doi.org/10.3409/azc.58_2.181, 2015.
Navarro, N., Montuire, S., Laffont, R., Steimetz, E., Onofrei, C., and Royer, A.: Identifying Past Remains of Morphologically Similar Vole Species Using Molar Shapes, Quaternary, 1, 20, https://doi.org/10.3390/quat1030020, 2018.
Pearce, E. A., Mazier, F., Normand, S., Fyfe, R., Andrieu, V., Bakels, C., Balwierz, Z., Binka, K., Boreham, S., Borisova, O. K., Brostrom, A., de Beaulieu, J.-L., Gao, C., González-Sampériz, P., Granoszewki, W., Hrynowizcka, A., Kolaczek, P., Kunes, P., Magri, D., Malkiewcz, M., Mighall, T., Milner, A. M., Möller, P., Nita, M., Noryskiewics, B., Agnieszka Pidel, I., Reille, M., Robertsson, A.-M., Salonen, J. S., Schläfli, P. Schokker, J., Scussolini, P., Seiriené, V., Strahl, J., Urban, B., Winter, H., and Svenning, J. C.: Substantial light woodland and open vegetation characterized the temperate forest biome before Homo sapiens, Sci. Adv., 9, https://doi.org/10.1126/sciadv.adi9135, 2023.
Pelletier, M., Royer, A., Holliday, T. W., Discamps, E., Madelaine, S., and Maureille, B.: Rabbits in the grave! Consequences of bioturbation on the Neandertal “burial” at Regourdou (Montignac-sur-Vézère, Dordogne), J. Hum. Evol., 110, 1–17, https://doi.org/10.1016/j.jhevol.2017.04.001, 2017.
Peltier, W. R., Argus, D. F., and Drummond, R.: Space geodesy constrains ice-age terminal deglaciation: The global ICE-6G_C (VM5a) model, J. Geophys. Res.-Sol. Ea., 120, 450–487, https://doi.org/10.1002/2014JB011176, 2015.
Pérez-Crespo, V. A., Rodríguez, J., Arroyo Cabrales, J., and Alva Valdivia, L. M.: Variación ambiental durante el Pleistoceno tardío y Holoceno temprano en Guilá Naquitz (Oaxaca, México), Rev. Bras. Paleontol., 16, 487–494, https://doi.org/10.4072/rbp.2013.3.08, 2013.
Perpinan Lamigueiro, O., Hijmans R., and Courtiol, A.: rasterVis, R package version 0.51.6 [code], https://oscarperpinan.codeberg.page/rastervis/ (last access: 15 October 2025), 2023.
Peyron, O., Guiot, J., Cheddadi, R., Tarasov, P., Reille, M., de Beaulieu, J. L., Bottema, S., and Andrieu, V.: Climatic reconstruction in Europe for 18,000 yr BP from pollen data, Quaternary Res., 49, 183–196, https://doi.org/10.1006/qres.1997.1961, 1998.
Peyron, O., Bégeot, C., Brewer, S., Heiri, O., Magny, M., Millet, L., Ruffaldi, P., Van Campo, E., and Yu, G.: Late-Glacial climatic changes in Eastern France (Lake Lautrey) from pollen, lake-levels, and chironomids, Quaternary Res., 64, 197–211, https://doi.org/10.1016/j.yqres.2005.01.006, 2005.
Pineda-Munoz, S., Wang, Y., Lyons, S. K., Tóth, A. B., and McGuire, J. L.: Mammal species occupy different climates following the expansion of human impacts, P. Natl. Acad. Sci. USA, 118, e1922859118, https://doi.org/10.1073/pnas.1922859118, 2021.
Piñero, P., Agustí, J., Blain, H.-A., and Laplana, C.: Paleoenvironmental reconstruction of the Early Pleistocene site of Quibas (SE Spain) using a rodent assemblage reconstruction, C. R.-Palevol., 15, 659–668, https://doi.org/10.1016/j.crpv.2015.06.009, 2016.
Piñero, P., López-García, J. M., Blain, H. A., Carnevale, G., Furió, M., Giuntelli, P., Luzi, E., Macaluso, L., Marramà, G., Pal, S., Pavia, G., Pavia, M., Pezzetti, C., Rocca, M., Sánchez-Bandera, C., Villa, A., and Delfino, M.: Multiproxy approach to reconstruct the climate and environment of a new late Middle Pleistocene vertebrate site in northwestern Italy, Palaeogeogr. Palaeoclimatol. Palaeoecol., 634, 111935, https://doi.org/10.1016/j.palaeo.2023.111935, 2024.
Ponomarev, D., van Kolfschoten, T., and van der Plicht, J.: Late Glacial and Holocene small mammals of the Timan Ridge (Komi Republic, Russia), Quatern. Int., 284, 177–183, https://doi.org/10.1016/j.quaint.2012.04.027, 2013.
Ponomarev, D., van Kolfschoten, T., van der Plicht, J., and Kosintsev, P.: Lateglacial desman discovered in Sed'yu-1 (Komi Republic, Russia), a site in the far northeast of Europe, Quatern. Int., 378, 88–98, https://doi.org/10.1016/j.quaint.2014.03.001, 2015.
Ponomarev, D. V., van Kolfschoten, T., and van der Plicht, J.: Late Glacial and Holocene micromammals of northeastern Europe, Russ. J. Theriol., 11, 121–130, 2012.
Prentice, I. C., Harrison, S. P., Jolly, D., and Guiot, J.: The climate and biomes of Europe at 6000 yr BP: comparison of model simulations and pollen-based reconstructions, Quaternary Sci. Rev., 17, 659–668, https://doi.org/10.1016/S0277-3791(98)00016-X., 1998.
Price, C. R.: Late Pleistocene and Early Holocene Small mammals in South West Britain, BAR Bristish series 347, Archaeopress, 105 pp., ISBN: 10 1841714852, 2003.
Puzachenko, A. Y. and Markova, A. K.: Evolution of mammal species composition and species richness during the Late Pleistocene-Holocene transition in Europe: A general view at the regional scale, Quatern. Int., 530, 88–106, https://doi.org/10.1016/j.quaint.2018.12.025, 2019.
Puzachenko, A. Y. and Markova, A. K.: The Scandinavian ice sheet against the Atlantic ocean: How the Scandinavian ice sheet affected European small mammal assemblage during the Greenland stadial GS-2.1, Quaternary Sci. Rev., 305, 108013, https://doi.org/10.1016/j.quascirev.2023.108013., 2023.
Quéré, J. P. and Le Louarn, H.: Les rongeurs de France: Faunistique et biologie, 3e édition revue et augmentée, Editions Quae, 311 pp., ISBN-13: 978-2759210336, 2011.
R Core Team: R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing, Vienna, Austria, https://www.R-project.org/ (last access: 15 October 2025), 2023.
Randall, D. A.: General circulation model development: past, present, and future, Elsevier, 416 pp., ISBN: 9780080507231, 2000.
Rasmussen, S. O., Bigler, M., Blockley, S. P., Blunier, T., Buchardt, S. L., Clausen, H. B., Cvijanovic, I., Dahl-Jensen ,D., Johnsen S. J., Fischer, H., Gkinis, V., Guillevic M., Hoek, W. Z., Lowe, J. J., Pedro, J. B., Popp, T., Seierstad, I. K., Steffensen, J. P., Svensson A. M., Vallelonga P., Vinther, B. M., Walker, M. J. C., Wheatley, J. J., and Winstrup, M.: A stratigraphic framework for abrupt climatic changes during the Last Glacial period based on three synchronized Greenland ice-core records: refining and extending the INTIMATE event stratigraphy, Quaternary Sci. Rev., 106, 14–28, https://doi.org/10.1016/j.quascirev.2014.09.007, 2014.
Reimer, P. J., Austin, W. E.N, Bard, E., Bayliss, A., Blackwell, P. G., Bronk Ramsey, C., Butzin, M., Cheng, H., Edwards, R. L., Friedrich, M., Grootes, P. M., Guilderson, T. P., Hajdas, I., Heaton, T. J., Hogg, A. G., Hughen, K. A., Kromer, B., Manning, S. W., Muscheler, R., Palmer, J. G., Pearson, C., van der Plicht, J., Reimer, R. W., Richards, D. A., Scott, E. M., Southon, J. R., Turney, C. S. M., Wacker, L., Adolphi, F., Büntgen, U., Capano, M., Fahrni, S. M., Fogtmann-Schulz, A., Friedrich, R., Köhler, P., Kudsk, S., Miyake, F., Olsen, J., Reinig, F., Sakamoto, M., Sookdeo, A., and Talamo, S.: The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP), Radiocarbon, 62, 725–757, https://doi.org/10.1017/RDC.2020.41, 2020.
Rey-Rodríguez, I., Gamarra, B., Arnaud, J., Golovanov, S., Kandel, A. W., Gasparyan, B., Wilkinson, K. N., Adler, D. S., and Weissbrod, L.: Climatic variability in the Armenian Highlands as the backdrop to hominin population dynamics 50–25 ka, Palaeogeogr. Palaeoclimatol. Palaeoecol., 112285, https://doi.org/10.1016/j.palaeo.2024.112285, 2024.
Rofes, J., García-Ibaibarriaga, N., Aguirre, M., Martínez-García, B., Ortega, L., Zuluaga, M. C., Bailon, S., Alonso-Olazabal, A., Castaños, J., and Murelaga, X.: Combining small-vertebrate, marine and stable-isotope data to reconstruct past environments, Sci. Rep.-UK, 5, 14219, https://doi.org/10.1038/srep14219, 2015.
Rofes, J., Cersoy, S., Zazzo, A., Royer, A., Nicod, P. Y., Laroulandie, V., Langlais, M., Pailler, Y., Leandri, C., Lebon, M., and Tresset, A.: Detecting stratigraphical issues using direct radiocarbon dating from small-mammal remains, J. Quaternary Sci., 35, 505–513, https://doi.org/10.1002/jqs.3193, 2020.
Rousset, F., Ferdy, J. B., Courtiol, A., and Eddelbuettel, D.: spaMM: mixed models, particularly spatial GLMMs, R package version [code], 1, https://doi.org/10.32614/CRAN.package.spaMM, 2016.
Royer, A.: Intérêts de l'utilisation des décapages lors des fouilles archéologiques pour l'étude des restes de petits vertébrés, PALEO, 25, 277–286, https://doi.org/10.4000/paleo.2819, 2014.
Royer, A.: How complex is the evolution of small mammal communities during the Late Glacial in southwest France?, Quatern. Int., 414, 23–33, https://doi.org/10.1016/j.quaint.2015.12.065, 2016.
Royer, A.: AurelienRoyer/Climate-spatial-Interpolations-LGM-LG: version submitted on 21.02.2025, Version V1, Zenodo [code], https://doi.org/10.5281/zenodo.14905209, 2025.
Royer, A., Lécuyer, C., Montuire, S., Escarguel, G., Fourel, F., Mann, A., and Maureille, B.: Late Pleistocene (MIS 3–4) climate inferred from micromammal communities and δ18O of rodents from Les Pradelles, France, Quaternary Res., 80, 113–124, https://doi.org/10.1016/j.yqres.2013.03.007, 2013.
Royer, A., Lécuyer, C., Montuire, S., Primault, J., Fourel, F., and Jeannet, M.: Summer air temperature, reconstructions from the last glacial stage based on rodents from the site Taillis-des-Coteaux (Vienne), Western France, Quaternary Res., 82, 420–429, https://doi.org/10.1016/j.yqres.2014.06.006, 2014.
Royer, A., Montuire, S., Legendre, S., Discamps, E., Jeannet, M., and Lécuyer, C.: Investigating the influence of climate changes on rodent communities at a regional-scale (MIS 1-3, Southwestern France), PLOS ONE, 11, e0145600, https://doi.org/10.1371/journal.pone.0145600, 2016.
Royer, A., Sécher, A., and Langlais, M.: A Brief Note on the Presence of the Common Hamster during the Late Glacial Period in Southwestern France, Quaternary, 1, 8, https://doi.org/10.3390/quat1010008, 2018.
Royer, A., Garcia Yelo, B. A., Laffont, R., and Hernandez Fernandez, M.: New bioclimatic models for the Quaternary Palaearctic based on insectivore and rodent communities, Palaeogeogr. Palaeoclimatol. Palaeoecol., 560, 110040, https://doi.org/10.1016/j.palaeo.2020.110040, 2020.
Royer A., Laroulandie V. Bailon S., Boudadi-Maligne M., Costamagno S., Danger M., Mallye J. B., and Rofes J.: Chapitre 3. Des restes de faune aux paléoenvironnements de Peyrazet, in: La Grotte-abri de Peyrazet (Creysse, Lot, France) au Magdalénien: Originalité fonctionnelle d'un habitat des derniers chasseurs de rennes du Quercy, edited by: Langlais, M. and Laroulandie, V., CNRS, Supplément à Gallia Préhistoire, 43, 35–48, ISBN: 978-2-271-13647-3, 2021.
Salzmann, U., Haywood, A. M., Lunt, D. J., Valdes, P. J., and Hill, D. J.: A new global biome reconstruction and data-model comparison for the Middle Pliocene, Glob. Ecol. Biogeogr., 17, 432–447, https://doi.org/10.1111/j.1466-8238.2008.00381.x., 2008.
Schürch, B., Wong, G. L., Luzi, E., and Conard, N. J.: Evidence for an earlier Magdalenian presence in the Lone Valley of southwest Germany, Journal of Archaeological Science: Reports, 57, 104632, https://doi.org/10.1016/j.jasrep.2024.104632, 2024.
Sesé, C.: Interpretación paleoclimática de las faunas de micromamíferos del Mioceno, Plioceno y Pleistoceno de la Cuenca de Guadix-Baza (Granada, España), Estud. Geol., 47, 73–83, https://doi.org/10.3989/egeol.91471-2409, 1991.
Simard-Pelissier, S.: Étude paléontologique des micromammifères de la grotte du Régourdou, PHD thesis, Faculté des Sciences de l'Université de Paris, 78 p., 1966.
Sloan, L. C., Crowley, T. J., and Pollard, D.: Modeling of middle Pliocene climate with the NCAR GENESIS general circulation model, Mar. Micropaleontol., 27, 51–61, https://doi.org/10.1016/0377-8398(95)00063-1, 1996.
Socha, P.: Rodent palaeofaunas from Biśnik Cave (Kraków-Częstochowa Upland, Poland): palaeoecological, palaeoclimatic and biostratigraphic reconstruction, Quatern. Int., 326, 64–81, https://doi.org/10.1016/j.quaint.2013.12.027, 2014.
Stewart, J. R. and Parfitt, S. A.: Late Quaternary environmental change at Walou Cave: evidence from a preliminary analysis of the small mammals, in: La Grotte Walou à Trooz (Belgique). Fouilles 1996 à 2004, edited by: Draily, C., Pirson, S., and Toussaint, M., Département du Patrimoine et l'Institut du Patrimoine Wallon, Namur, 38–59, ISBN-13 978-2875220455, 2011.
Stewart, J. R., Lister, A. M., Barnes, I., and Dalén, L.: Refugia revisited: individualistic responses of species in space and time, Proc. Biol. Sci. B, 277, 661–671, https://doi.org/10.1098/rspb.2009.1272, 2010.
Stoetzel, E., Lalis, A., Nicolas, V., Aulagnier, S., Benazzou, T., Dauphin, Y., Abdeljalil El Hajraoui, M., El Hassani, A., Fahd, S., Fekhaou M., Geigle, E.-V., Lapointe, F.-J., Leblois, R., Ohler, A., Nespoulet, R., and Denys, C.: Quaternary terrestrial microvertebrates from mediterranean northwestern Africa: State-of-the-art focused on recent multidisciplinary studies, Quaternary Sci. Rev., 224, 105966, https://doi.org/10.1016/j.quascirev.2019.105966, 2019.
Stoetzel, E., Rodrigues, H. G., and Cornette, R.: Towards a better knowledge of the molar morphology and ecology of extant and fossil grass rats (Muridae: Arvicanthis Lesson, 1842), Quaternary Res., 114, 1–16, https://doi.org/10.1017/qua.2023.7, 2023.
Stoetzel, E., Lebreton, L., Pallier, C., Bruxelles, L., Bon, F., Lejay, M., Anderson, L., Martin, H., and Jarry, M.: ?The small vertebrate assemblages from the Aurignacian deposits of the Mas d'Azil cave (Ariège, France): new data for the understanding of the MIS3 environmental changes in the Pyrenees, C. R.-Palevol., 2025, 1–18, https://doi.org/10.5852/cr-palevol2025v24a1, 2025.
Sueyoshi, T., Ohgaito, R., Yamamoto, A., Chikamoto, M. O., Hajima, T., Okajima, H., Yoshimori, M., Abe, M., O'ishi, R., Saito, F., Watanabe, S., Kawamiya, M., and Abe-Ouchi, A.: Set-up of the PMIP3 paleoclimate experiments conducted using an Earth system model, MIROC-ESM, Geosci. Model Dev., 6, 819–836, https://doi.org/10.5194/gmd-6-819-2013, 2013.
Surov, A., Banaszek, A., Bogomolov, P., Feoktistova, N., and Monecke, S.: Dramatic global decrease in the range and reproduction rate of the European hamster Cricetus cricetus, Endanger. Species Res., 31, 119–145, https://doi.org/10.3354/esr00749, 2016.
Tarasov, P. E., Volkova, V. S., Webb III, T., Guiot, J., Andreev, A. A., Bezusko, L. G., Bykova, G. V., Dorofeyuk N. I., Kvavadze E. V., Osipova I. M., Panova N. K., and Sevastyanov, D. V.: Last glacial maximum biomes reconstructed from pollen and plant macrofossil data from northern Eurasia, J. Biogeogr., 27, 609–620, https://doi.org/10.1046/j.1365-2699.2000.00429.x., 2000.
Taylor, K. E., Stouffer, R. J., and Meehl, G. A.: An overview of CMIP5 and the experiment design, B. Am. Meteorol. Soc, 93, 485–498, https://doi.org/10.1175/BAMS-D-11-00094.1, 2012.
Tchernov, E.: Rodent faunas and environmental changes in the Pleistocene of Israel, in: Rodents in desert environments, edited by: Prakash, I., Springer, Dordrecht, 331–362, ISBN: 978-94-010-1946-0, 1975.
Terray, L., Stoetzel, E., Ben Arous, E., Kageyama, M., Cornette, R., and Braconnot, P.: Refinement of the environmental and chronological context of the archeological site El Harhoura 2 (Rabat, Morocco) using paleoclimatic simulations, Clim. Past, 19, 1245–1263, https://doi.org/10.5194/cp-19-1245-2023, 2023.
Teterina, A.: Rodents of the North Urals in the Late Pleistocene and Holocene, Quatern. Int., 201, 31–36, https://doi.org/10.1016/j.quaint.2008.05.018, 2009.
Tougard, C., Renvoisé, E., Petitjean, A., and Quéré, J. P.: New insight into the colonization processes of common voles: inferences from molecular and fossil evidence, PLoS One, 3, e3532, https://doi.org/10.1371/journal.pone.0003532, 2008.
van Dam, J. A.: Geographic and temporal patterns in the late Neogene (12–3 Ma) aridification of Europe: the use of small mammals as paleoprecipitation proxies, Palaeogeogr. Palaeoclimatol. Palaeoecol., 238, 190–218, https://doi.org/10.1016/j.palaeo.2006.03.025, 2006.
van de Weerd, A. and Daams, R.: Quantitative composition of rodent faunas in the Spanish Neogene and paleoecological implications (I and II), Proceedings of the Koninklijke Nederlandse Akademie Van Wetenschappen, Series B, 81, 448–473, 1978.
van den Hoek Ostende, L.: Insectivores (Lipotyphla, Mammalia) from the Ramblian of the Daroca-Calamocha area, Coloquios de paleontología, 281–310, ISSN 1132-1660, 2003.
van der Meulen, A. J. and Daams, R.: Evolution of Early-Middle Miocene rodent faunas in relation to long-term palaeoenvironmental changes, Palaeogeogr. Palaeoclimatol. Palaeoecol., 93, 227–253, https://doi.org/10.1016/0031-0182(92)90099-Q, 1992.
Varela, S., Lima-Ribeiro, M. S., and Terribile, L. C.: A short guide to the climatic variables of the last glacial maximum for biogeographers, PLOS ONE, 10, e0129037, https://doi.org/10.1371/journal.pone.0129037, 2015.
Voldoire, A., Sanchez-Gomez, E., Salas y Mélia, D., Decharme, B., Cassou, C., Sénési, S., Valcke, S., Beau, I., Alias, A., Chevallier, M., Déqué, M., Deshayes, J., Douville, H., Fernandez, E., Madec, G., Maisonnave, E., Moine, M.-P., Planton, S., Saint-Martin, D., Szopa, S., Tyteca, S., Alkama, R., Belamari, S., Braune, A., Coquart, L., and Chauvin, F.: The CNRM-CM5. 1 global climate model: description and basic evaluation, Clim. Dynam., 40, 2091–2121, https://doi.org/10.1007/s00382-011-1259-y, 2013.
Walter, H.: Vegetationszonen und Klima, Eugen Ulmer, Stuttgart, ISBN-13: 978-3800124015, 1970.
Williams, J. W. and Jackson, S. T.: Novel climates, no-analog communities, and ecological surprises, Front. Ecol. Environ., 5, 475–482, https://doi.org/10.1890/070037, 2007.
Wilson, D. E. and Mittermeier Jr., R. A.: The Mammals of the World-Vol. 8, Insectivores, Sloths and Colugos, Lynx Edicions, Barcelona, 709 pp., ISBN-13: 978-8416728084, 2018.
Wilson, D. E., Lacher, T. E., and Mittermeier Jr., R. A.: The Mammals of the World-Vol. 7, Rodents II, Lynx Edicions, Barcelona, 1008 pp., ISBN-13: 978-8416728046, 2017.
Wolff, J. O. and Sherman, P. W.: Rodent societies: an ecological and evolutionary perspective, University of Chicago Press, 610 pp., ISBN-13: 978-0226905372, 2008.
Wong, G. L., Starkovich, B. M., and Conard, N. J.: Human subsistence and environment during the Magdalenian at Langmahdhalde: evidence from a new rock shelter in the Lone Valley, southwest Germany, Mitteilungen der Gesellschaft für Urgeschichte, 26, 103–123, 2017.
Wong, G. L., Drucker, D. G., Starkovich, B. M., and Conard, N. J.: Latest Pleistocene paleoenvironmental reconstructions from the Swabian Jura, southwestern Germany: evidence from stable isotope analysis and micromammal remains, Palaeogeogr. Palaeoclimatol. Palaeoecol., 540, 109527, https://doi.org/10.1016/j.palaeo.2019.109527, 2020.
Woodman, P., McCarthy, M., and Monaghan, N.: The Irish quaternary fauna project, Quaternary Sci. Rev., 16, 129–159, https://doi.org/10.1016/S0277-3791(96)00037-6, 1997.
Wu, H., Guiot, J., Brewer, S., and Guo, Z.: Climatic changes in Eurasia and Africa at the last glacial maximum and mid-Holocene: reconstruction from pollen data using inverse vegetation modelling, Clim. Dynam., 29, 211–229, https://doi.org/10.1007/s00382-007-0231-3, 2007.
Yukimoto, S., Adachi, Y., Hosaka, M., Sakami, T., Yoshimura, H., Hirabara, M., Tanaka, T. Y., Shindo, E., Tsujino, H., Deushi, M., Mizuta, R., Yabu, S., Obata, A., Nakano, H., Koshiro, T., Ose, T., and Kitoh, A.: A new global climate model of the Meteorological Research Institute: MRI-CGCM3 – Model description and basic performance, J. Meteorol. Soc. Jpn Ser. II, 90A, 23–64, https://doi.org/10.2151/jmsj.2012-A02, 2012.
Zheng, W. and Yu, Y.: Paleoclimate simulations of the mid-Holocene and Last Glacial Maximum by FGOALS, Adv. Atmos. Sci., 30, 684–698, https://doi.org/10.1007/s00376-012-2177-6, 2013.